Нейронный контроль движений глаз
Контроль над положением глаз и координация сокращения наружных мышц глаза интенсивно изучались и изучаются до настоящего времени при помощи клинических, анатомических, физиологических и биоинженерных методов исследования. Интеграция данных этих дисциплин способствовала пониманию деятель-
Движения глаз
 ности глазодвигательной системы животных и человека.
ности глазодвигательной системы животных и человека.
Основными функциями нейронного контроля движений глаза является обеспечение направления глаза к зрительной цели, поддержание функции бинокулярного зрения, пространственная локализация зрительной цели. Происходит это благодаря доставке центростремительной информации коре головного мозга сведений относительно положения глаза в глазнице, что, в свою очередь, помогает определить направление взора [17, 83, 96, 201, 527].
Афферентная система. Система, регулирующая движения глаза, состоит из приводящей части (афферентная система), передающей сенсорную информацию относительно положения глаза, а также отводящей части (эфферентная система), ответственной за его движение [414, 498].
Чувствительные волокна, идущие от наружных мышц глаза в составе III, IV и VI нервов, направляются к нейронам, расположенным в ядре среднемозгового тракта тройничного нерва [17, 96, 201, 424, 498, 554]. Конечным пунктом передачи информации являются чувствительные ядра тройничного нерва, а таже вентральное основное ядро зрительного бугра [96, 201, 306, 360, 528].
Информация к перечисленным структурам нервной системы поступает от многочисленных механорецепторов, залегающих в паренхиме наружных мышц глаза.
К основным механорецепторам относятся: мышечное веретено, сухожильный аппарат Гольджи, палисадоподобные и спиральные нервные окончания. Выявляются и другие рецеп-торные структуры, отличающиеся от аналогичных образований скелетной мускулатуры как по строению, так и по распределению в мышце (табл. 4.4.3).
Мышечное веретено. В наружных мышцах глаза мышечное веретено обнаружено как у животных, так и у человека [90, ПО, 122, 367, 376, 510]. Мышечное веретено наружных мышц глаза отличается от таковых скелетной мускулатуры [474].
Основной функцией этой структуры является точная регистрация положения глазного яблока в глазнице и передача этой информации вышерасположенным нервным центрам [62, 346, 465].
Мышечное веретено распределено неравномерно и чаще обнаруживается в узкой части мышцы. Число этих рецепторов в одной мышце колеблется от 22 до 71. Внутренняя прямая мышца глаза содержит 18,8 мышечного веретена; наружная прямая—19,3; нижняя прямая — 34,0; верхняя косая — 27,3; нижняя косая — 4,3.
Каждое веретено содержит в капсуле 1 — 15 интрафузальных волокон [376, 465], а на полюсе обнаруживается концевая пластинка. В об-
Таблица 4.4.3. Чувствительный аппарат наружных мышц глаза человека
| Мышечный рецептор | Особенности распределения |
| и строения | |
| Классическое мышеч- | В передней и задней трети |
| ное веретено | мышцы |
| Атипичное мышечное | Инкапсулированные нервные |
| веретено | волокна |
| Сухожильный аппарат | Схожи с аппаратом Гольджи |
| Гольджи | скелетной мышцы и распола- |
| гаются в сухожилии | |
| Палисадообразные | Инкапсулированные нервные |
| окончания | окончания в месте мышеч- |
| но-сухожильного соединения | |
| Спиральные окончания: | Неинкапсулированные |
| Одиночные спираль- | Расположены в средней тре- |
| ные окончания | ти мышцы |
| Двойные спираль- | Неинкапсулированные нерв- |
| ные окончания | ные волокна. Обычно множе- |
| ство ответвлений | |
| Множественные | Множество «спиралей» от |
| спиральные оконча- | многочисленных нервных во- |
| ния | локон |
| Другие рецепторы: | Обнаруживаются в соедини- |
| Подобные цветку | тельной ткани. Встречаются |
| пластинки | редко и их строение не впол- |
| Древоподобные чув- | не исследовано |
| ствительные окон- | |
| чания | |
| Щеткоподобные чув- | |
| ствительные оконча- | |
| ния | |
| Катушкоподобные | |
| чувствительные | |
| структуры | |
| Луковицеподобные | |
| чувствительные | |
| окончания |
ласти экватора рецептора располагается множество мякотных и безмякотных нервных волокон, окружающих интрафузальное волокно. Толщина интрафузальных волокон различна и колеблется от 5 до 30 мкм (в среднем 10 мкм). Длина веретена равняется 350—725 мкм (в среднем — 500 мкм) [457, 471].
Рядом исследователей установлено, что мышечное веретено не регистрирует положение глазного яблока при отсутствии зрительного стимула [73]. Fuchs, Kornhuber [171] предполагают, что веретено поставляет информацию мозжечку относительно длины и конечного пункта саккадического движения глаза.
Сухожильный аппарат Гольджи регистрирует степень регидности мышцы [465, 523]. Длина каждой отдельной структуры равна примерно 1 мм, а ширина — 0,1 мм. В каждом образовании выявляется 15—20 экстрафузаль-ных волокон. Капсула состоит из сплетения коллагеновых волокон, между которыми проходят приводящие аксоны.
Глава 4.ГОЛОВНОЙ МОЗГ И ГЛАЗ
 Сокращение мышцы сжимает аксоны, в результате чего формируются сигналы, свидетельствующие об увеличении ригидности мышцы.
Сокращение мышцы сжимает аксоны, в результате чего формируются сигналы, свидетельствующие об увеличении ригидности мышцы.
Палисадоподобные окончания. Палисадо-подобные окончания (мышечно-сухожильные цилиндры) являются основным сенсорным аппаратом наружных мышц глаза млекопитающих [347]. Они состоят из «скрытых» нервных окончаний, находящихся в месте перехода мышечной ткани в сухожилие. Эти образования представляют собой нежную сеть нервных волокон, контактирующих с мышечным волокном [441].Один мякотный аксон может отдавать ветви нескольким соседним мышечным волокнам. В большем количестве они обнаружены в горизонтальных прямых мышцах глаза [25, 77, 458, 459, 475, 523]. Нервный сигнал формируется в результате сжатия терминалов при сокращении мышцы.
Спиральные» окончания. В средней трети наружных мышц глаза выявляются нервные волокна, спиралевидно окутывающие мышечные волокна. Они делают 3—8 поворотов вокруг мышечного волокна и заканчиваются структурой, напоминающей концевую пластинку.
| III II И II I II I N III II I I I I II II III II I I I I I * "»\ 11 11 I I I I I I I I |

|
Тракты проприорецепции.Проприорецеп-тивные импульсы от наружных мышц глаза проходят через глазную ветвь тройничного нерва к полулунному ганглию, обладающему соматотопической организацией (рис. 4.4.11)

В
1 MM
| Illl | —— | • • • • |
| ВнП |

 НрП Врк~ВрП НК—НП
НрП Врк~ВрП НК—НП
Рис. 4.4.11. Диаграмма, иллюстрирующая соматотопи-
ческую организацию афферентов проприорецептивной
чувствительности наружных мышц глаза в полулунном
ганглии {no E. Manni, Е. V. Petrossi [354]):
Л — латеральная; Д — дорзальная; М — медиальная; В— вентральная поверхности полулунного ганглия; НрП—наружная прямая мышца; ВрК — верхняя косая мышца; ВрП—верхняя прямая мышца; НК—нижняя косая мышца; НП — нижняя прямая мышца; ВнП—внутренняя прямая мышца
[354]. Центральные отростки нейронов первого порядка заканчиваются в ипсилатеральном чувствительном ядре тройничного нерва. Они также проецируются на более каудальную часть ядра тройничного нерва [414].
От ядра тройничного нерва проприорецеп-тивные импульсы передаются крыше и покрышке среднего мозга и посредством медиальной петли {lemniscus medialis) поступают к вентрально расположенному комплексу ядер зрительного бугра [498]. К другим путям, вовлеченным в анализ проприорецептивной информации, относятся наружное коленчатое тело, зрительная кора головного мозга [77, 414]. Проприорецептивные волокна от наружных мышц глаза подходят также к V, VI и VII долям коры мозжечка посредством мшистых волокон [354—359, 424]. Участвуют в обработке информации и верхние холмики четверохолмия [14, 33, 84, 147]. При этом проприорецептивные афференты к верхнему четверохолмию обладают тормозным влиянием.
Эфферентная система.Цель глазодвигательной системы состоит в том, чтобы навести глаза на выбранный объект наиболее эффективным и скоординированным способом. В предыдущих главах нами показано, что степень остроты зрения, в определенной мере, определяется плотностью расположения фоторецепторов в специализированной части сетчатой оболочки, называемой центральной ямкой. Естественно, что в эволюционном развитии параллельно с формированием центрального зрения развивалась и система наведения изображения именно на этот наиболее чувствительный участок сетчатки. Раньше развились системы, позволяющие фиксировать неподвижное изображение. Подобный тип фиксации взора характерен для низших позвоночных. У более сложно организованных животных появилась необходимость следить за движущимися целями и при этом фиксировать изображение на наиболее чувствительный участок сетчатой оболочки. Подобный тип фиксации взора характерен для высших млекопитающих, включая человека. Благодаря столь сложному эволюционному развитию системы слежения за зрительной целью и сформировалось пять систем, функционирующих содружественно и дополняющих одна другую. Это саккадная система, система плавного слежения, вергентная система, система обеспечения фиксации и вестибулярная система. Фунционирование первых четырех систем зависят от информации, получаемой от органа зрения, в то время как формирование последней (вестибулярной) зависит от информации, получаемой от вестибулярных и отоли-товых рецепторов, рецепторов шеи и, возможно, от наружных мышц глаза. Информация от всех перечисленных систем объединяется, обрабатывается и направляется к ядрам, управляющим наружными мышцами глаза (табл. 4.4.4).
Движения глаз
 Таблица 4.4.4. Характеристика различных типов движения глаз
Таблица 4.4.4. Характеристика различных типов движения глаз
| Поддерживающее | Плавное слежение | Саккады | Вергентные дви- | Вестибулярный | |
| положение | жения | рефлекс | |||
| Функция | Поддерживает | Поддерживает | Быстро переме- | Выравнивание | Установление |
| направление гла- | положение дви- | щает цель на | зрительной оси | положения глаз | |
| за в направлении | жущейся цели | фовеа | для достижения | при изменении | |
| цели | на фовеа | фовеолярной | положения голо- | ||
| фиксации | вы и туловища | ||||
| Стимулы | «Зрительный ин- | Движение объек- | Цель по перифе- | Диспарантность | Стимуляция по- |
| терес» и «внима- | та вблизи фовеа | рии поля зрения | сетчаток | лукружных ка- | |
| ние» | налов, мешочка | ||||
| и маточки | |||||
| Латентный | — | 125 мс | 200 мс | 160 мс | Очень короткий |
| период | |||||
| (время от | |||||
| стимула | |||||
| до движе- | |||||
| ния глаз) | |||||
| Скорость | Большая (резкие | До 100° в с | До 400° в с | Около 20° в с | До 300° в с |
| движения, микро- | (обычно 30° | ||||
| саккады и медлен- | ВС) | ||||
| ные — «дрейф») | |||||
| Обратная | __ | Продолжитель- | Отобранная ин- | __ | |
| связь | ная | формация | |||
| Субстрат | Затылочно-темен- | Затылочно-темен- | Лобная, затылоч- | Неизвестен | Вестибулярный |
| ная кора | ная кора | но-теменная кора, | аппарат, мышеч- | ||
| верхние бугор- | ные рецепторы | ||||
| ки четверохолмия | шеи, мозжечок |
Саккады. Саккадная система обеспечивает обнаружение зрительной цели и выведение ее на наиболее чувствительную часть сетчатой оболочки (центральная ямка), исправляя при этом существующую «угловую ошибку» между ними. При формировании саккад используется зрительная, соматосенсорная и слуховая информация. Информация о величине и направлении необходимого для фиксации изображения на сетчатке изменения положения глаза («двигательная ошибка») передается стволу мозга, в котором и вычисляются статические и динамические силы, необходимые для выполнения этого движения [96, 201].
Саккады являются быстрыми скачкоподоб-ными движениями глаза из одной точки фиксации к другой. Они чередуются с периодами фиксации, длящимися примерно 0,15—2,0 с. Амплитуда этих движений колеблется от нескольких угловых минут до нескольких градусов. Средняя угловая скорость при этом тем выше, чем больше амплитуда саккады, и доходит до 200—600° в секунду. Длительность саккады варьирует от 10 до 80 мс. Саккады с амплитудой 10—15° обычно сопровождаются дополнительным поворотом головы.
Типичное саккадическое движение глаза у человека равно десяти градусам и продолжается 45 мс. При произвольных саккадах длительность ее равняется 200 мс. Непроизвольные саккады более быстрые. Они бывают во сне, а также в течение быстрой стадии оптокинетического и вестибулярного нистагма, во время
зрительной моторной реакции при неожиданном появлении цели по периферии поля зрения. Нередко в процессе саккадического движения глазное яблоко как бы «промахивается» относительно цели, т. е. цель не фиксируется в центральной ямке, а располагается дальше. Саккадное «промахивание» (дисметрическая саккада) встречается даже в норме. После дис-метрической саккады всегда наступает корректирующее движение. В норме встречается два типа корректирующих движений: корректирующая саккада с определенным интервалом времени ожидания между наступившей и предыдущей саккадами и корректирующее «скользящее» движение, развивающееся без интервала при завершении оригинала саккады. «Скользящее» движение не требует нового управляющего импульса, идущего от мозга.
Саккадические движения нередко нарушаются при различных заболеваниях, сопровождающихся поражением головного мозга и глаза [219]. Саккады отсутствуют при тяжелой прогрессивной надъядерной офтальмоплегии, при поражении лобно-среднемозговых (мезенцефа-лических) трактов. Нарушение временных характеристик саккад, их скорости и точности обнаруживается у больных с рассеянным склерозом в 57%. К нарушению саккадических движений может привести применение ряда фармакологических препаратов [187].
Анатомический субстрат саккад. Традиционно считают, что генерирует саккадические движения кора лобной области головного мозга
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 после поступления в нее необходимой информации от органа зрения и наружных мышц глаза. Это добавочные поля первичной лобной зрительной коры, включая дополнительную зрительную кору и дорзальную прелобную кору. Контролируют саккады также поле 7а затылочной коры, теменная кора, латеральное внут-ризатылочное поле, наружное коленчатое тело и базальные ганглии [17, 96, 201, 364, 394, 395]. Все вышеперечисленные области проецируются на верхние бугорки четверохолмия (рис. 4.4.12, 4.4.13). Существуют данные относительно того, что перечисленные области мозга формируют сигнал для «пакетных нейронов». Участие и взаимодействие того или иного участка коры головного мозга зависит от типа саккадического движения, что проиллюстрировано рис. 4.4.12, 4.4.13 и табл. 4.4.5.
после поступления в нее необходимой информации от органа зрения и наружных мышц глаза. Это добавочные поля первичной лобной зрительной коры, включая дополнительную зрительную кору и дорзальную прелобную кору. Контролируют саккады также поле 7а затылочной коры, теменная кора, латеральное внут-ризатылочное поле, наружное коленчатое тело и базальные ганглии [17, 96, 201, 364, 394, 395]. Все вышеперечисленные области проецируются на верхние бугорки четверохолмия (рис. 4.4.12, 4.4.13). Существуют данные относительно того, что перечисленные области мозга формируют сигнал для «пакетных нейронов». Участие и взаимодействие того или иного участка коры головного мозга зависит от типа саккадического движения, что проиллюстрировано рис. 4.4.12, 4.4.13 и табл. 4.4.5.
Наиболее важной областью формирования саккад является кора лобной глазной области. В отличие от теменной коры и базальных ганглиев, кора лобной глазной области проецируется непосредственно на центры движения глаз, расположенные в мосту [96, 201]. После повреждения верхних бугорков четверохолмия только лобная кора способна генерировать саккады.
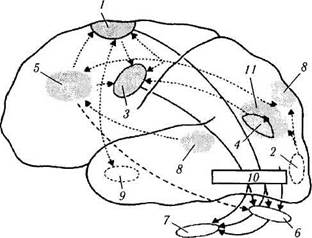
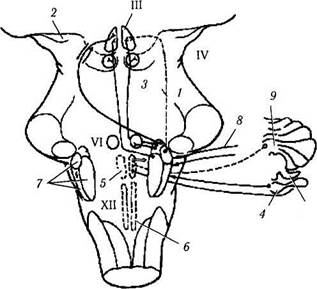
Рис. 4.4.12. Гипотетическая схема взаимодействия различных участков коры головного мозга в процессе саккадического движения глаз (по Pierrot—Deseilligny, Ann Neurol 1995, [37, 557—567]):
/—дополнительное глазодвигательное поле; 2 — затылочная кора; 3—лобное глазодвигательное поле; 4—теменное глазодвигательное поле; 5 — прелобная кора; 6 — верхние бугорки четверохолмия; 7—парамедианная ретикулярная формация варольева моста; 8—вестибулярная кора; 9—гиппокамп; 10 — зрительный бугор; // — теменная задняя кора


|
| МПП |
| О |
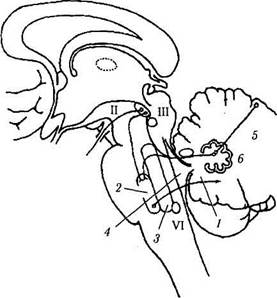
Рис. 4.4.13. Обобщенная схема нейрального контроля движений глазных яблок (no Miller, 1985):
рисунок в центре показывает связь между глазной лобной областью (ГЛО) и теменно-височно-затылочной (ТВЗ) областью с верхними бугорками четверохолмия (ВБ), а также ростральным промежуточным ядром медиального продольного пучка (рпМПП) и парамедианной ретикулярной формацией варольева моста {ПРФМ). ГЛО и ВБ продуцируют саккады, в то время как ТВЗ является наиболее важным участком коры в контроле плавного слежения. Схематическое изображение, приведенное слева, иллюстрирует тракты в области ствола мозга, контролирующие горизонтальный взгляд. Аксоны клеточных тел, локализованных в ПРМФ, подходят к ипсилатеральному ядру отводящего нерва (VI), где они образуют синапс с мото-
нейронами отводящего нерва, аксоны которых, перекрещиваясь по средней линии, направляются в составе медиального продольного пучка (МПП) к части ядра глазодвигательного нерва, иннервирующей внутреннюю прямую мышцу (ВП) противоположной стороны. Схематическое изображение справа иллюстрирует тракт ствола мозга вертикального взгляда. К наиболее важным структурам относятся рпМПП, ПРФМ, промежуточное ядро Кахала (ЯК) и задняя спайка (ЗС). Аксоны нейронов, расположенных в вестибулярном ядре (ВЯ), проходят мимо ядра отводящего нерва и посредством медиального продольного пучка подходят к ядрам глазодвигательного (III) и блокового (IV) нервов
Авижения глаз
Таблица 4.4.5. Взаимодействие различных участков коры головного мозга в формировании саккадических
движений различного типа
 ЗБ
ЗБ
ВК
ЗК
ТЗК
тгп
ПК
ЛГП
дпг
г
ВБ
 Рефлекторные саккады, вызванные зрительным стимулом
Рефлекторные саккады, вызванные зрительным стимулом
Ингибирование саккад, вызванных зрительным стимулом
Программирование саккад, вызванные стимулом, исходящим не из сетчатой оболочки
Саккады «памяти» в результате зрительного сигнала
Саккады «памяти» в результате вестибулярного сигнала
Последовательные саккады «памяти», вызванные зрительным сигналом
Примечания. ЗБ— зрительный бугор; ВК— вестибулярная кора; ЗК—затылочная кора; ТЗК—теменная зрительная кора; ЛГП — лобное глазодвигательное поле; 7777 — теменное глазодвигательное поле; ПК — пре-лобная кора; ДПГ — дополнительное глазодвигательное поле; Г — гиппокамп; ВБ — верхние бугорки четверохолмия. Саккада «памяти» представляет собой повторную саккаду, при которой используется информация, полученная во время первой саккады.
| К наружной прямой мышце |
Как выше указано, нисходящий тракт от лобной коры сводится к прямой проекции на мост, а именно на его парамедианную ретикулярную формацию [240]. Существует и непрямая проекция. При этом образуются синапсы в наружном коленчатом теле, и только затем проецируется на ретикулярную формацию (рис. 4.4.12, 4.4.13). Парамедианная ретикулярная формация играет решающую роль в генерации глазных движений. Располагается она между глазодвигательными ядрами по срединной линии ствола мозга. Предполагают, что парамедианная ретикулярная формация в физиологическом отношении состоит из двух частей. Большая часть расположена рострально. Функционирует она как «генератор импульсов», поскольку в эксперименте обнаружены в этой области нейроны, формирующие «пакет» потенциалов действия,
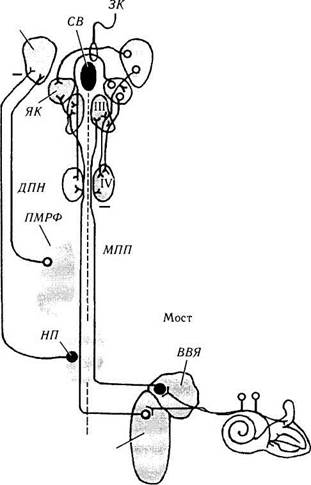
 Рис. 4.4.14. Схематическое изображение генератора саккадических движений глаз, расположенного в стволе мозга (no Albert, 1995):
Рис. 4.4.14. Схематическое изображение генератора саккадических движений глаз, расположенного в стволе мозга (no Albert, 1995):
изменение положения глаза (Е) и картина активности нейронов при саккадическом движении в левую сторону. Вход к стволу мозга происходит от верхних бугорков четверохолмия (ВБ) и глазного поля лобной коры (ГПЛК). Эти сигналы «перекрещиваются» в стволе мозга и возбуждают «пакетные» («физические») нейроны контрлатеральной парамедианной ретикулярной формации моста (ПМРФ). Длительно функционирующие «пакетные» нейроны (ДПН) разряжаются и возбуждают нейроны, функционирующие со средней длительностью (СПН). Именно эти нейроны кодируют так называемый «скоростной сигнал» саккад. Нейроны «паузы» (НП) расположены в вентрокаудальной части парамедианной ретикулярной формации в nucleus raphe inter-positus (ядро шва — ЯШ). Эти клетки тормозят «пакетные» нейроны, функционирующие только с целью генерации саккад. Тормозные «пакетные» нейроны (77/) лежат каудальней ядра отводящего нерва (VI) в дорзолатеральной части ретикулярной формации продолговатого мозга (ПРФПМ) и тормозят контрлатеральные мотонейроны. Сигнал о положении глаза генерируется «нейральным интегратором» в медиальном вестибулярном ядре (МВЯ) и nucleus prepositus hypoglossy. Разряжение мотонейронов (МН) кодирует как скорость саккадического движения, так и его амплитуду («шаг»). Заштрихованные кружки — тормозные нейроны, а полые — возбуждающие
начинающихся приблизительно за 8—10 мс перед движением глаза [188, 298]. Эти нейроны названы «пакетными» нейронами (рис. 4.4.14). Выявлено также, что число импульсов этих нейронов линейно связано с амплитудой саккады в горизонтальной плоскости.
Каудальная часть парамедианной ретикулярной формации, лежащая вблизи ядер отводящего нерва, выполняет функцию «нейраль-ного интегратора» и содержит «тонические» нейроны и «нейроны паузы» [240, 298, 339, 383] (рис. 4.1.14). Располагаются клетки «ней-
| Генератор импульсов |
| ДПН |
| СПН |
| К наружной прямой мышце |
| Интегратор |
| ПРФПМ |
 ГПЛК
ГПЛК
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
рального интегратора» в nucleus prepositus hypoglossi и медиальном вестибулярном ядре. Нейроны этого ядра получают входы от пре-моторных «саккадных полей» ствола мозга и проецируются на ядра глазодвигательных нервов, мозжечок и вестибулярные ядра [565] (рис. 4.4.14).
Подобное взаимодействие позволяет клеткам «нейрального интегратора» объединять получаемую сенсорную информацию и формировать сигнал, определяющий частоту и амплитуду саккады. При этом амплитуда саккад кодируется в премоторных центрах, обеспеченных топографическими сенсорными картами стриар-ной коры, лобной глазной области и верхних бугорков четверохолмия [96, 201, 395].
Особое место занимают верхние бугорки четверохолмия. Еще в конце XIX века выявлено, что стимуляция верхних бугорков приводит к возникновению саккадоподобных движений глаз. В 70-е годы XX столетия на основании экспериментальных электорофизиологических исследований установлены важные факты. Во-первых, верхние бугорки не генерируют саккады. Они определяют точность движения глаз по направлению к цели, а также определяют направление и амплитуду движения независимо от первоначального положения глаз в орбите, поскольку содержат «топографическую карту» движений глаза [96, 201, 279, 280, 426, 525, 611]. Подтверждается это тем, что стимуляция соседних участков бугорков приводит к смещению взгляда к смежным точкам цели. Во-вторых, верхние бугорки четверохолмия участвуют и в координации движений глаза в ответ на звуковой раздражитель, соматосенсорную информацию, обеспечивая, таким образом, направление взгляда в сторону звукового сигнала или тактильного раздражителя.
Теперь мы несколько подробней остановимся на нейронном контроле саккад в горизонтальной плоскости. Формируется премоторная команда горизонтальной саккады, как и других движений в горизонтальной плоскости, в нейронах парамедианной ретикулярной формации моста [240, 243, 385], а затем проецируется на ипсилатеральное ядро отводящего нерва (рис. 4.4.13—4.4.15).
Ядро отводящего нерва лежит непосредственно по срединной линии в дорзальной части моста, сразу под дном четвертого желудочка. Оно содержит два типа нейронов. Большие мотонейроны ядра отдают аксоны, которые проходят вентрально через мост и выходят на внутренней поверхности ствола мозга, иннервируя наружную прямую мышцу глаза [535].
Ядро содержит также другие, маленькие, нейроны, чьи аксоны не покидают ствол мозга, а, перекрещиваясь, переходят на противоположную сторону, где входят в состав медиального продольного пучка и заканчиваются на нейронах ядер глазодвигательного нерва
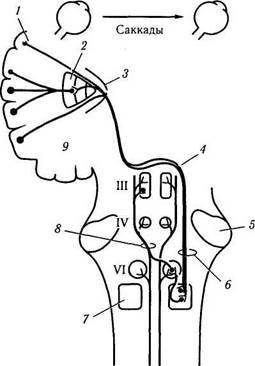
Рис. 4.4.15. Надъядерный контроль саккадических
движений глаза, горизонтального взгляда в правую
сторону (цель расположена в контрлатеральном поле)
(по Reeh et al., 1985):
I — лобное глазное двигательное поле (8); 2 — базальные ганглии; 3—внутренняя капсула; 4 — среднемозговой (мезанцефа-лический) перекрест; 5 — ножка мозжечка; 6—кортикобульбар-ный тракт (кортикотегментальный тракт); 7 — парамедианная ретикулярная формация варольева моста; 8—медиальный продольный пучок; 9—дополнительное двигательное поле (7). Основной путь показан красным цветом, а дополнительный путь, который может модифицировать горизонтальный взгляд, показан черным цветом
(рис. 4.4.14, 4.4.15). Именно благодаря этой связи и возбуждаются мотонейроны внутренней прямой мышцы глаза противоположной стороны, сокращение которой переводит взгляд.
«Тонические» нейроны ядра отводящего нерва возбуждаются перед началом саккадическо-го движения и сохраняют такое состояние на протяжении всего движения.
Тормозные «тонические» нейроны располагаются несколько каудальней ядра отводящего нерва и обеспечивают взаимную иннервацию, посылая тормозные сигналы к контрлатеральному отводящему ядру [209, 241, 271]. Такая нейронная структура отвечает принципу антагонистической иннервации.
Вертикальные саккады также регулируются нейронами парамедианной ретикулярной формации моста [240]. Но связь между этими нейронами и глазодвигательными ядрами не является прямой, а осуществляется через ретикулярную формацию среднего мозга. Кроме того, формируют вертикальный взгляд промежуточное (интерстициальное) ядро медиального продольного пучка, лежащее вблизи
глаз

|
| РяМПП |
| Средний мозг |
| I 1 МВЯ |
| Лабиринт |
зрительного бугра с вентральной его стороны, а также ядро Кахала [88, 96, 201, 383]. Нейроны интерстициального ядра проецируются на нейроны глазодвигательного ядра, контролирующие верхнюю и нижнюю прямые мышцы и нижнюю косую мышцу. Более подробно ней-
Продолговатый мозг
Рис. 4.4.16. Нейронный контроль содружественных вертикальных движений глаз:
вестибулярные афференты от передних и задних полукружных каналов проецируются на верхнее (ВВЯ) и медиальное (МВЯ) вестибулярные ядра. Проекции от вестибулярных нейронов поднимаются в составе медиального продольного пучка (МПП) и соединительного плеча (не показано) к мотонейронам блокового нерва (IV), а также к ядру глазодвигательного нерва (III) и ядру Кахала (ЯК). Вестибулярные проекции, проходящие в ипсилате-ральном МПП, ингибированы и исходят из верхнего вестибулярного ядра. Аксоны длительно функционирующих «пакетных» нейрнов (ДПН) восходят из парамедианной ретикулярной формации моста (ПМРФ) и направляются к ростральному промежуточному (интерстициальному) ядру медиального продольного пучка (ряМПП). Нейроны последнего кодируют вертикальные саккады и быструю стадию нистагма. Восходящие аксоны перекрещиваются несколько ростральней сильвиевого водопровода (СВ) в задней комиссуре (ЗК), в то время как нейроны, иннервиру-ющие мотонейроны депрессора, проходят несколько латераль-ней и через ядро Кахала. Ядро Кахала и его реципроктные связи с вестибулярными ядрами формируют «нейральный интегратор» вертикальных движений. Команды плавного слежения проецируются на клочок (флоккулус) мозжечка от вестибулярного ядра. Заштрихованные кружки — тормозные нейроны, полые — возбуждающие; НП — нейроны паузы
ронная связь между ядрами представлена на рис. 4.4.13 и 4.4.16.
Необходимо отметить, что нейроны парамедианной ретикулярной формации моста возбуждаются за несколько миллисекунд до начала вертикальной саккады. При этом они «кодируют» направление саккад. На эти клетки проецируются на верхние бугорки четверохолмия, кору лобной области и ядро клиновидного пучка (п. cuneatus), лежащее с латеральной стороны ретикулярной формации среднего мозга [114,240, 573].
Нейроны ядра шва (п. raphes), расположенные около ядра отводящего нерва, тормозят «тонические» клетки между саккадами и в момент фиксации. Если эти нейроны, называемые нейронами «паузы», стимулировать во время саккады, движение глаза прерывается [296]. Новое положение глаза в конце саккад поддерживается потоком импульсов, являющихся результатом функционирования «тонических» нейронов.
Плавное слежение.Плавные (следящие) движения глаз необходимы для длительной фиксации движущегося объекта и слежения за ним. При этом угловая скорость вращения глаз примерно соответствует скорости движения цели, если скорость последней не превышает 60—80° в секунду. Наиболее точно система плавного слежения работает тогда, когда скорость перемещения цели ниже 30° в секунду. После того как цель выбрана, система работает вне волевого контроля [96, 201, 556].
На плавные движения глаз накладываются коррекционные саккады. В этом случае изображение фиксируемой точки предмета остается в пределах 2° от центральной ямки. При скорости перемещения зрительного объекта более 80° в секунду следящие движения глаз происходят значительно медленнее, чем движение предмета. По этой причине изображение передвигается по сетчатке. Если скорость движения изображения не превышает 180° в секунду, то смещение изображения по сетчатке компенсируется дополнительными саккадическими движениями большей амплитуды. Координация таких следящих движений глаз обычно осуществляется за счет последующих движений головы. Координация движений головы и глаз обычно осуществляется в центрах управления взгляда, расположенных в стволе мозга. Информация относительно положения головы и положения глаз поступает в эти центры не только от вестибулярных рецепторов внутреннего уха, но и от механорецепторов суставов шеи.
Вышеперечисленные сложные функции слежения, сочетающиеся с саккадами и находящиеся под контролем вестибулярной системы и мозжечка, обеспечиваются сложной системой нервного контроля, о чем подробно будет изложено ниже.
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 | |
|
| Q |
| Q |
| Плавные движения |
Анатомический субстрат плавного слежения. Надъядерный контроль плавного слежения пока точно не выяснен. На рис. 4.4.17— 4.4.19 приведены лишь упрощенные схемы. От контроля саккад контроль плавного слежения отличается тем, что проекция от коры головного мозга на парамедианную ретикулярную формацию осуществляется с этой же стороны.
Предполагают, что сигнал, индуцированный зрительным стимулом, возникает в затылочной коре головного мозга. Нейроны коры этой области проецируются на среднее височное зрительное поле. В свою очередь, нейроны среднего височного зрительного поля проецируются на медиальное верхнее височное зрительное поле, оказывающее помощь в слежении за зрительной целью в тех случаях, когда она временно исчезает из поля зрения. Обе височные проекции, в свою очередь, проецируются на затылочную кору. В плавном слежении участвуют также теменная и лобная зрительная кора [141, 543].
Нисходящий путь от коры головного мозга направляется с этой же стороны к ретикулярной формации моста, латеральным ядрам моста и латеральным ядрам добавочной зрительной системы (рис. 4.4.17—4.4.19). Добавочная зрительная система, в свою очередь, проецируется на мозжечок, который затем проецируется на медиальные вестибулярные ядра. Конечной точкой проекции являются ядра глазодвигательных нервов [123]. При этом волокна дважды перекрещиваются (в среднем мозге — перекрест Мейнерта (Meynert) и в области моста).
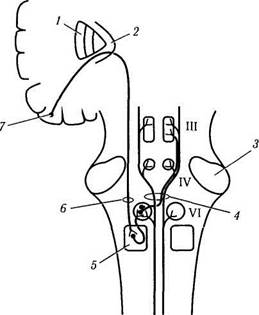
Рис. 4.4.17. Надъядерный контроль плавного слежения
в горизонтальной плоскости в левую сторону (цель
появляется в контрлатеральном поле) (по Reek et al.,
1981):
I—базальные ганглии; 2— внутренняя капсула; 3 — ножка мозжечка; 4 — медиальный продольный пучок; 5 — парамедиан-ная ретикулярная формация варольева моста; 6—кортико-бульбарный тракт; 7 — ассоциативные поля затылочной коры. Основной путь обозначен красным цветом, остальные — черным
 Рис. 4.4.18. Схема связей зрительного тракта с верхними бугорками четверохолмия, подушкой зрительного бугра и «добавочной зрительной системой»:
Рис. 4.4.18. Схема связей зрительного тракта с верхними бугорками четверохолмия, подушкой зрительного бугра и «добавочной зрительной системой»:
/ — нейроны сетчатки; 2 — кора; 3 — наружное коленчатое тело; 4 — зрительный перекрест; 5 — подушка зрительного бугра; б — верхние бугорки четверохолмия; 7—претектальная область; 8—добавочная зрительная система
Движения глаз

|
|
| 10 |
а 6
Рис. 4.4.19. Связи мозжечка, определяющие движения глаз (по Bron et al., 1997):
а — со стволовой частью мозга (/ — нижняя мозжечковая ножка; 2 — ретикулярное ядро крыши моста; 3— парамедианная ретикулярная формация моста; 4— верхняя мозжечковая ножка; 5 — полушарие; 6— червь; II, III, VI — ядра черепно-мозговых нервов); б — с вестибулярными ядрами (/—добавочный тракт Дейтерса; 2— соединительное плечо; 3— медиальный продольный пучок; 4 — узелок мозжечка; 5 — necl. prepositus hypoglossi; 6 — nucl. perihypoglossi; 7 — вестибулярные ядра; 8 — первичные вестибулярные афференты; 9 — червь; 10 — язычок червя)
Добавочная зрительная система состоит из скопления нейронов, расположенных вне первичной зрительной коры и получающих проекции от сетчатки (рис. 4.4.18). К ней относятся претектальные ядра, в состав которых входит ядро зрительного тракта, ядра крыши моста, медиальное ядро моста, п. perihypoglossi, вестибулярная комиссура.
Основной функцией добавочной зрительной системы является интеграция получаемой зрительной информации, а также информации о положении тела и головы в пространстве, получаемой от вестибулярной, соматосенсорной и зрительной систем. Благодаря этой интеграции получаемой информации и формируются сигналы, идущие к исполнительным нервным центрам, участвующим в движении глаза [596—603].
В контроле горизонтальных саккадических движений, как указано выше, принимают участие ретикулярная формация среднего мозга и ряд структур мозжечка. Ретикулярная формация среднего мозга (formatio reticularis) содержит нейроны, «кодирующие» положение глаза. Некоторые из них активны в течение ипси-латерального плавного слежения или в течение медленной стадии вестибулярного рефлекса. Другие нейроны, находящиеся ниже ядра отводящего нерва, формируют премоторную команду, определяющую соответствие скорости движения глаз скорости слежения за целью [298].
Мозжечок является ключевой структурой в функционировании плавного слежения. Струк-
турой, соединяющей кору головного мозга, глазодвигательную часть мозжечка и ствол мозга, является дорзолатеральное ядро ствола мозга. Это ядро содержит нейроны, кодирующие направление и скорость движения цели и глаза и координируют их. Проецируется дорзо-медиальное ядро ствола мозга, в первую очередь, на мозжечок.
Клочок мозжечка (flocculus) получает зрительную информацию посредством мшистых волокон. Клетки Пуркине (определяющие скорость слежения) клочка суммируют сигналы, поступающие от вестибулярной и зрительной системы [17, 530]. При этом они формируют сигналы, определяющие скорость движения глаза за целью. Клетки Пуркине проецируются на п. perihypoglossi или парамедианную ретикулярную формацию моста, т. е. структуры, важные в заключительном синтезе команды, определяющей направление взгляда [96, 201, 337, 338, 377] (рис. 4.4.19).
Зубчатое ядро (nucleus dentatus) и лежащая вблизи Y-образная группа вестибулярных ядер (вероятно, получающая проекции от клочка) вовлечены в контроль вертикального плавного слежения и отмены вестибуло-глазного рефлекса [372].
Система плавного слежения нередко повреждается, причем раньше, чем саккады. Наступает это при двухстороннем поражении ба-зальных ганглиев и ствола мозга. Подобные состояния характерны для алкогольного и бар-битуратного отравления. При этом скорость
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 плавного слежения снижается и больной начинает использовать саккады для контроля цели даже при ее небольшой скорости движения.
плавного слежения снижается и больной начинает использовать саккады для контроля цели даже при ее небольшой скорости движения.
Система, контролирующая фиксацию глаз.Фиксация взгляда на неподвижную цель длится, по крайней мере, пять секунд. При этом глазное яблоко, тем не менее, находится в движении. Эти движения глаза включают быстрые движения (микросаккады) и медленные клони-ческие движения.
Микросаккады никогда не прекращаются. Их амплитуда всего несколько угловых секунд, а частота находится в переделах 20—150 Гц.
В результате медленного клонического «дрейфа» точка фиксации уходит от фиксируемого объекта. С помощью микросаккад происходит компенсация этих дрейфов и восстановление правильной фиксации объекта.
Вергентные движения.Вергентные движения относятся к так называемым версиям. Отличие сводится к тому, что при версионных движениях зрительные оси движущихся глаз остаются параллельными, а при вергентных угол между ними изменяется [96, 201, 518].
Способностью вергентных движений обладает только небольшое число видов позвоночных и приматы. Функцией вергентных движений является достижение проекции изображения на область центральной ямки каждого глаза. Это достигается управлением изменения угла между зрительными осями двух глаз.
Различают три источника стимула начала вергентных движений. Первый стимул — это так называемое «бинокулярное неравенство».
Понятие «бинокулярное неравенство» характеризует состояние, при котором изображение проецируется на сетчатку двух глаз, но на различные ее участки. Вторым стимулом является нерезкость (стертость) изображения, связанная с отсутствием адекватной аккомодации. По всей видимости, состояние хрусталиков обоих глаз контролируется высшими центрами нервной системы и используется для вычисления расстояния до зрительной цели. Третий стимул, относящийся к так называемым «тоническим», учитывает состояние системы при отсутствии конвергенции, что бывает, например, в условиях полной темноты. У человека в подобных условиях угол конвергенции равен 3°. Последний тип информации, используемый в вергентных движениях, — это информация о монокулярной линейной глубине (линейная перспектива), определяющая расстояние до цели в монокулярных условиях.
Вергентные движения глаз относятся к медленным движениям. Система вступает в действие при смещении точки фиксации в пространстве. При этом направление движения одного глаза является зеркальным отображением движения другого. Если фиксируемая точка на значительном расстоянии от наблюдателя, то зрительные оси параллельны. Если взор смещается на более близкий предмет, зрительные оси конвергируют (рис. 4.4.20). При необходимости перевода взора на отдаленный предмет оси разводятся (дивергенция). В обычных условиях акту конвергенции осей глаза сопутствует сужение зрачка и аккомодация [412]. Эти три компонента формируют так называемый «кон-
|

|

|
 А
А
Вблизи
I
Вдали
i > «
T
Цель 1
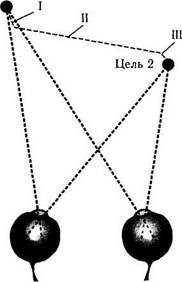
Рис. 4.4.20. Вергентные движения глаз:
а — изменение угла конвергенции в зависимости от расстояния до цели; б — характеристика траектории саккадического движения глаз при смещении цели вправо и ближе к наблюдателю. Вергеитный механизм срабатывает раньше конвергенции (I). Затем начинается саккада и степень конвергенции усиливается (II). После прекращения саккадического движения вергентный механизм продолжает действовать до тех пор, пока цель не будет выведена на фовеолы обоих глаз (III)
Движения глаз
 вергентно-аккомодационно-зрачковый рефлекс» (аккомодационный рефлекс).
вергентно-аккомодационно-зрачковый рефлекс» (аккомодационный рефлекс).
Вергентная система начинает работать после саккадического движения, спустя i римерно 160 мс. Скорость движения глаз при этом равняется 20° в секунду. Конвергенция или дивергенция зрительных осей происходит со скоростью 10° в секунду. Это медленное движение может длиться почти секунду.
Анатомический субстрат вергентных движений. Надъядерный контроль вергентной системы изучен далеко не достаточно.
Известно, что высшие зрительные центры коры головного мозга особо важны в восприятии неравенства изображений на сетчатках и инициирования вергентных движений глаза. Об этом свидетельствует то, что одновременная стимуляция зрительных полей лобной и затылочной коры приводит к конвергенции (стимуляция полей 19 и 22 коры затылочной доли). На диспарантность сетчаток реагируют также поля коры средней височной и затылочной областей. Промежуточная премоторная инициация вергенции определяется также стимуляцией нейронов, расположенных вблизи ядер глазодвигательного нерва, которые проецируются на ядро внутренней прямой мышцы глаза и ядро отводящего нерва [186, 2Э1, 518].
Выявлены еще другие участки мозга, участвующие в индукции вергентных движений. Это — средний мозг, мост. Установлено, что участок среднего мозга, лежащий латеральней глазодвигательного ядра, контролирует угол конвергенции. К вергентным движениям приводит также стимуляция моста между медиальными продольными пучками. В вергентных движениях участвует и мозжечок, о чем свидетельствует тот факт, что его ампутация приводит к нарушению движения глаз.
В последние годы появились данные о том, что «центром вергентных движений» являются скопления двух типов нейронов ретикулярной формации моста [186, 240, 370, 379]. По своим функциям он напоминает «центр саккад». Первый тип нейронов, «пакетные» нейроны, формирует импульсы перед началом вергентного движения. При этом частота импульсов связана со скоростью движения, а их количество — с амплитудой. Вовлечены при этом «пакетные» нейроны, обеспечивающие как схождение, так и расхождение зрительных осей. «Тонические вергентные нейроны» объединяют динамические и статические реакции вергентной системы. Необходимо отметить и то, что вергентные движения интимно связаны с саккадами. Эта связь проявляется рядом этапов совместных действий.
Рефлекторные системы, не связанные со зрительной системой (вестибулярная).Рефлекторные изменения движения глаз происходят всякий раз, когда изменяется положение головы и шеи [77]. При эт'ом возбуждаются
рецепторы полукружных каналов или макуляр-ных органов. Эти рефлекторные движения глаз осуществляются за счет прямых связей между нейронами вестибулярных ядер, мотонейронами глазных мышц и нейронами парамедианной ретикулярной формации моста и ретикулярной формации среднего мозга.
Сигналы от рецепторов шеи, вестибулярного аппарата и от наружных мышц глаза определяют положение головы относительно тела и в пространстве, а также положение глазных яблок. Таким образом, формируется «эфферентная копия», обходящая зрительный путь.
Вестибулярная система может компенсировать значительно более существенные движения изображения на сетчатке, чем система плавного слежения. Поворот туловища является стимулом компенсационных движений глаза, контролируемых вестибулярной системой. Скорость движений глаза, индуцированных вестибулярной системой, достигает до 300° в секунду. Движение головы более 10—15° приводит к саккадическому движению глаз в противоположном направлении.
Несмотря на то, что вестибулярная система не контролируется зрительной системой, все же между ними существует взаимодействие, называемое вестибуло-глазным рефлексом [372].
Движения глаза в вестибуло-глазной системе содружественные и состоят из двух стадий. Первая стадия — это плавное следящее движение. Прерывается оно саккадами, которые повторно приводят глаза к средней позиции в глазнице. Этот тип повторяющихся медленных и быстрых движений глаза, относится к одному из типов нистагма (вестибулярный нистагм).
Необходимо помнить то, что тип нистагма определяется направлением быстрой стадии. Так, например, левосторонний нистагм характеризуется быстрым движением влево. Только медленная стадия нистагма компенсирует движения туловища.
Нистагм может быть индуцирован обычными физиологическими стимулами или являться выражением различных заболеваний, нарушающих нормальный контроль движений глаза.
Анатомический субстрат вестибулярного и оптокинетического рефлексов.Основной функцией вестибулярной системы и мозжечка является определение направления взора при любом положении и движении тела.
Перед тем как остановиться на рефлексах, нам представляется необходимым более подробно описать строение вестибулярного аппарата и мозжечка, т. е. структур, во многом определяющих возможность этой функции.
Первоначально необходимо охарактеризовать саму вестибулярную рефлекторную дугу.
Строение вестибулярного аппарата. Вкостном лабиринте височной кости лежат три (с каждой стороны) мембранных полукружных
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
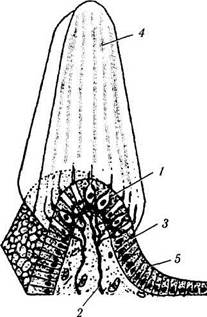
канала (canalis semicircularis anterior, posterior, lateralis), выполненных эндолимфой [4, 6—9, 11, 578]. Как показано на приведенных рисунках (рис. 4.4.21—4.4.23), существует один горизонтальный и два вертикальных канала, которые лежат в трех плоскостях, перпендикулярных друг другу. Горизонтальные (латеральные) каналы с двух сторон лежат в одной плоскости, в то время как плоскость переднего канала одной стороны параллельна плоскости заднего канала противоположной стороны. Горизонтальные каналы соединяются с одного конца с маточкой, которая, по сути, является большим расширением перепончатой части лабиринта. Вертикальные каналы (передний и задний) соединяются с маточкой своим одним концом, а другим концом соединяются друг с другом. Расширения одного конца полукружных каналов называются ампулой (ampula ossea anterior, posterior, lateralis). Каждая ампула содержит «волосковые» чувствительные клетки, погруженные в желатиноподобный материал, называемый капулой (рис. 4.4.22).
|
| 12 |
| 10 |
Таким образом, вестибулярный орган состоит из двух морфологических субъединиц, а именно: статолитового аппарата (macula utri-culi и macula sacculi) и полукружных каналов (передний и задний вертикальные и горизонтальный каналы). В области макул и в полу-
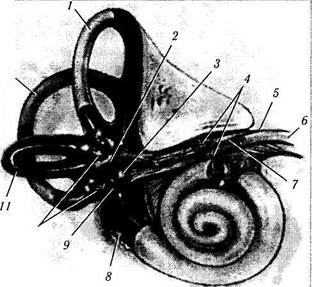
Рис. 4.4.21. Органы и нервы внутреннего уха (по Walsh, 1969):
1 — верхний полукружный канал; 2— маточка; 3 — мешочек; 4 — вестибулярный ганглий; 5 — внутренний слуховой проход; б — лицевой нерв; 7—вестибулярная ветвь VIII нерва; 8—круглое окно; 9 — овальное окно; 10 — ампульные нервы; // — горизонтальный полукружный канал; 12— задний полукружный канал. Вид спереди лабиринта и раковины (кохлеа). Обращает на себя внимание наличие тесных анатомических отношений между вестибулярной и кохлеарной ветвями пред-дверно-улиткового (VIII) и лицевого (VII) нервов во внутреннем слуховом канале. Также видны три ампулярных нерва (терминальные концы нервов на иллюстрации обрезаны), которые собирают информацию от каждого полукружного канала
кружных каналах вблизи от так называемых ампул расположен содержащий рецепторы сенсорный эпителий, покрытый желеобразной массой. В макулах эта масса покрывает сенсорные клетки и содержит соли кальция в виде кристаллов кальцита (отолитовая мембрана). В полукружных каналах желеобразная масса кристаллов не содержит.
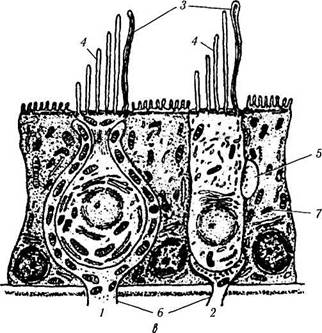
В вестибулярном органе имеется два типа клеток («волосковые» клетки), на апикальной поверхности которых располагаются реснички. Различают два типа ресничек — стереоцилии (по 60—80 ресничек в одной клетке) и. киноци-лии (по одной на каждой клетке) (рис. 4.4.22).
Рецепторы относятся к вторичным сенсорным клеткам. Как указано выше, они не обладают собственными отростками, а иннервиру-ются афферентными волокнами нейронов вестибулярного ганглия, входящими в состав вестибулярного нерва (рис. 4.4.21). На рецептор-ных клетках также оканчиваются эфферентные нервные волокна.
Афферентные волокна передают информацию об уровне возбуждения периферического органа в центральную нервную систему, а эфферентные волокна изменяют чувствительность рецепторов.
Как было показано выше, реснички рецеп-торных клеток входят в отолитовую мембрану. Поскольку она содержит соли кальция, ее удельный вес больше, чем эндолимфы, заполняющей полость мешочка (sacculus) и маточки (utriculus). По этой причине при действии на организм линейного ускорения отолитовый аппарат скользит по сенсорному эпителию. В результате этого реснички отклоняются и рецептор получает стимул.
Когда тело выпрямлено, а голова находится в нормальном положении, макула маточки оказывается под углом и тяжелая отолитовая мембрана соскальзывает по сенсорному эпителию. При этом реснички изгибаются и происходит стимуляция рецептора. В зависимости от направления наклона частота нервной импуль-сации в афферентных волокнах увеличивается или уменьшается.
Стимуляция макулы мешочка происходит так же, но эта структура расположена почти вертикально при нормальном положении головы. Таким образом, при любом положении головы каждая из отолитовых мембран занимает определенное положение относительно эпителия. При этом создается определенная картина возбуждения, отражающая расположение головы и тела в пространстве, которая затем обрабатывается в центральном отделе вестибулярной системы.
Второй вид адекватных стимулов для ресничек вестибулярных рецепторов воспринимается в полукружных каналах. Капула, расположенная в полукружных каналах, обладает такой же плотностью, что и эндолимфа. По этой причине
Движения глаз


|
|
|
Рис. 4.4.22. Строение вестибулярного аппарата:
а — распределение сенсорного эпителия в перепончатом лабиринте и улитке (темные области); б — схема строения вестибулярного аппарата млекопитающих (/ — эпителий перепончатого канала; 2 — отолиты; 3— отолитовая мембрана; 4 — поддерживающие клетки; 5 — волосковые клетки; 6 — нервные окончания); в — ультраструктура волосковых клеток органа равновесия (/ — волосковые клетки первого типа; 2— волосковые клетки второго типа; 3— стереоцилии; 4 — киноцилии; 5 — афферентные нервные окончания; 6 — эфферентные нервные окончания; 7 — поддерживающие клетки); г — ампулярный гребешок (/ — волосковые клетки; 2 — нервные волокна; 3 — поддерживающие клетки; 4 — купол; 5 — эпителий перепончатого канала)
линейное ускорение не влияет на этот орган. Возбуждение происходит при угловом ускорении (вращении). При повороте головы полукружные каналы поворачиваются вместе с ней. Однако эндолимфа в силу своей инерции в первый момент остается неподвижной. Поэтому капула отклоняется в направлении, противоположном направлению вращения. Это отклонение сдвигает реснички и таким образом изменяет активность в афферентном нерве. В горизонтальных каналах все рецепторы ориентированы
так, что киноцилии обращены к маточке, поэтому активность в афферентных волокнах увеличивается, когда капулы отклоняются в сторону маточки. В левом горизонтальном полукружном канале это происходит при вращении влево.
В вертикальных каналах активация нервных волокон происходит при утрикофугальном (в направлении, противоположном утрикулусу) отклонении капулы.
По картине активности нервных волокон, приходящих от трех полукружных каналов
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

| Поворот глаз |

 Плоскость расположения левого и правого латеральных каналов
Плоскость расположения левого и правого латеральных каналов
У
| Левый ПВК |
Правый ЗВК
Левый ЗВК
Правый
Н о
Q. О
ш о
С
Рис. 4.4.23. Плоскости расположения полукружных каналов с правой и левой стороны (a) (no Barber, Stockwell, 1976) и направление смещения эндолимфы при повороте головы в правую стороны (б) (по Cogan, 1966). Глаза
при этом поворачиваются влево:
ПВК—передний вертикальный канал; ЗВК — задний вертикальный канал
с каждой стороны, мозг получает информацию относительно углового ускорения головы [588].
Очевидно, что система наиболее чувствительна к вращениям, плоскость которых совпадает с плоскостью канала. Для «вычисления» скорости вращения головы в трех измерениях используются три отдельных канала, ориентированных в различных плоскостях. Фактически, имеется шесть каналов, по три с каждой стороны. Вращение головы в левую сторону активизирует два канала, расположенных в горизонтальной плоскости. «Волосковые» клетки в одном из этих каналов будут деполяризоваться, поскольку капула отклоняет их киноцилии. «Волосковые» клетки другого канала, наоборот, гиперполяризуются, так как капула отклоняется в противоположном направлении, отклоняя при этом киноцилии в против
Дата добавления: 2015-03-26; просмотров: 2030;