КЛЕТКА И ТКАНИ
ЗАДАЧЕЙ НАСТОЯЩЕЙ ГЛАВЫ является ознакомление читателя с современными сведениями о структурной организации клеток и тканей организма человека. Эти данные должны помочь легче воспринимать излагаемый ниже материал, особенно посвященный описанию микроскопического строения глаза, а также структурной организации нервных центров, обрабатывающих зрительную информацию и управляющих его движением. При этом мы исходили из того, что многие читатели довольно давно обращались к учебнику гистологии и не вполне четко помнят основные положения цитологии и гистологии. Необходимо учесть, что за время, истекшее с момента изучения читателем этого предмета, в представлениях о структуре клетки и ткани произошли довольно существенные изменения.
Данная глава основана на изложении материалов наиболее полных и современных монографий и учебников по гистологии [1—9]. При этом определенное внимание уделялось приведению сведений относительно структурного своеобразия тканевой организации глазного яблока и окружающих его образований.
1.1. КЛЕТКА
1.1.1. Цитоплазма и органоиды
Клетка является основной структурной единицей тканей и, естественно, органов и организма в целом.
В клетке четко различаются ядро и цитоплазма (рис. 1.1.1). Бесструктурная часть цито-
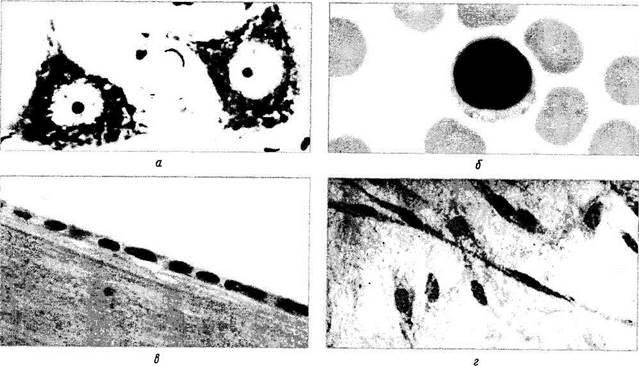
Рис. 1.1.1. Строение клеток различных тканей:
а — мотонейрон коры головного мозга; б — лимфоцит периферической крови и эритроциты; в — эпителий передней капсулы
хрусталика; г — фиброциты соединительной ткани
Глава 1. КЛЕТКА И ТКАНИ
 плазмы, выявляемая при световой микроскопии и расположенная по периферии клетки, называется эктоплазмой. Большая часть цитоплазмы обладает определенной структурой и называется эндоплазмой. Покрыта цитоплазма цито-плазматической мембраной (плазмолемма).
плазмы, выявляемая при световой микроскопии и расположенная по периферии клетки, называется эктоплазмой. Большая часть цитоплазмы обладает определенной структурой и называется эндоплазмой. Покрыта цитоплазма цито-плазматической мембраной (плазмолемма).
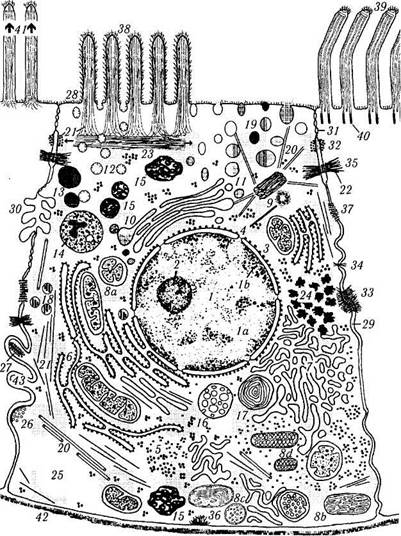
Ультраструктурные исследования выявили довольно сложное строение клетки (рис. 1.1.2, табл. 1.1.1).
Перед тем как остановиться на структурной организации клетки, необходимо описать строение биологической мембраны.
Биологическая мембрана. Каждая клетка окружена плазматической мембраной (плазмо-леммой), которую невозможно различить в световом микроскопе. Лишь при приготовлении ультратонких срезов плазматическая мембрана видна в виде трехслойной структуры (два элек-тронноплотных слоя, между которыми располагается светлый слой) (рис. 1.1.3). Общая толщина плазмолеммы равняется 7,5 нм, а толщина каждого из трех слоев приближается к 2,5 нм. Основа плазмолеммы — так называемая элементарная мембрана, являющаяся основ-
|
Рис. 1.1.2. Ультраструктурная организация клетки:
/ — ядро (1а— эухроматин; 1Ь— гетерохроматин); 2— ядрышко; 3 — ядерная мембрана; 4 — пора ядерной оболочки; 5 — рибосомы; 6 — шероховатый эндоплазматический ретикулум; 7 — гладкий эндоплазматический ретикулум; 8а — митохондрии, содержащие кристы; 8Ь—митохондрии тубулярного типа; 8с— митохондрии призматического типа; 8d — митохондрии мешочко-подобного типа; 9 — центриоли; 10— комплекс Гольджи; // — пузырьки комплекса Гольджи; 12 — эндоцитоплазматический пузырек (пиносома); 13— лизосома; 14 — вторичная лизосо-ма (аутофаголизосома); /5 — третичная лизосома (телолизо-сома, липофусциновая гранула); 16 — мультивезикулярное тельце; 17 — пластинчатое тело; 18 — пероксисома (микротельце);
19 — секреторная гранула; 20 — микротрубочки; 21 — актино-вые филаменты; 22 — десмосома; 23 — терминальная сеть; 24 — гранула гликогена; 25 — жировая капля; 26 — синапс; 27 — си-наптическая лента с пузырьками; 28 — клеточная мембрана с гликокаликсом; 29 — межклеточное пространство; 30 — инвагинация мембраны; 31 — плотное соединение (zonula occlu-dens); 32 — zonula adhaerens; 33 — fascia adhaerens; 34 — punctum adhaerens; 35 — macula adhaerens; 36 — полудесмо-сома; 37 — щелевое соединение (нексус); 38 — микроворсинки с гликокаликсом; 39 — реснички; 40 — базальное тельце реснички; 41—стереоцилии; 42—базальная пластинка; 43—интер-дигитации
Клетка
 Таблица 1.1.1. Структурные компоненты клетки
Таблица 1.1.1. Структурные компоненты клетки
| Плазмолемма | Цитоплазма | Ядро |
| Слои плазмолеммы: | Гиалоплазма (цитозоль) | Ядерная оболочка (кариотека) |
| — наружный; | Органеллы | Наружная мембрана |
| — промежуточный; | Мембранные | Внутренняя мембрана |
| — внутренний | Комплекс Гольджи | Перинуклеарное пространство |
| Эндоплазматическая сеть | Комплекс поры | |
| Структуры, в образовании которых | Гранулярная (зернистая, шерохова- | Нуклеоплазма |
| участвует плазмолемма: | тая) | Ядерная ламина |
| — клеточные отростки; | Агранулярная (гладкая) | Фибриллярная (нитчатая) |
| — микроворсинки; | Фагосомы | Гранулярная (зернистая) |
| — реснички; | Лизосомы | Ядрышко: |
| — жгутики | Первичная | — главное; |
| Фаголизосома | — добавочное | |
| Межклеточные соединения: | Аутофагосома | Хромосомы |
| — простые; | Остаточное тельце | Хроматин |
| — зубчатые; | Мультивезикулярное тельце | Эухроматин |
| — пальцевидные; | Пероксисомы | Гетерохроматин |
| — сложные; | Окаймленные пузырьки | Тельце полового хроматина |
| — пятно сцепления (десмосома); | Меланосомы | Гранула хроматина |
| — поясок сцепления (лентовидная | Немембранные | Центросома кинетохор |
| десмосома); | Свободные рибосомы и полирибо- | |
| — полудесмосома; | сомы | |
| — запирающая зона (плотное со- | Клеточный центр | |
| единение); | Центриоли | |
| — щелевое соединение (нексус) | Цитоскелет (фибриллярные струк- | |
| туры, опорный аппарат клетки) | ||
| Микротрубочки | ||
| Промежуточные филаменты | ||
| Микрофиламенты | ||
| Микрофибриллы | ||
| Включения | ||
| Гранулы | ||
| Гликогена | ||
| Белковые | ||
| Пигментные | ||
| Меланиновые | ||
| Секреторные | ||
| Капли жира (адипосомы) | ||
| Кристаллоидные |
ной структурной единицей всех мембранных образований клетки (клеточной оболочки, ядерной оболочки, мембранных органоидов).
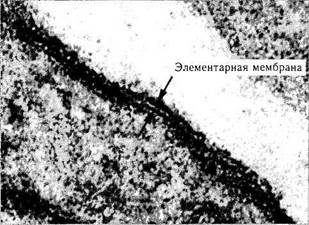
Рис. 1.1.3. Ультраструктурное строение элементарной мембраны
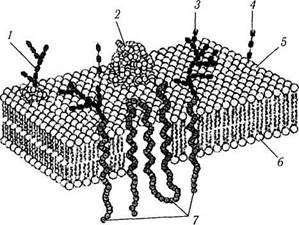
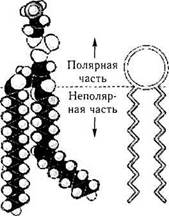
В химическом отношении элементарная мембрана представляет собой билипидный слой (рис. 1.1.4). Липиды, формирующие мембрану, в основном, относятся к фосфатидилхолину (лецитин) и фосфатидилэтаноламину (цефалин). Эти липиды отличаются полярностью. Один конец липидной молекулы гидрофобный, а противоположный— гидрофильный (рис. 1.1.4). Гидрофобные цепи обращены внутрь биослоя, а гидрофильные головки — к наружи. Именно благодаря этому и формируется двуслойность структуры. В большинство мембран входит также холестерин.
В билипидный слой плазмолеммы погружены молекулы белка, проходящие через всю толщину мембраны. Часть подобных глобулярных белковых телец образуют непрерывные белковые «каналы» между наружной и внутренней средой клетки. Белки, погруженные в липид-ный слой, называются интегральными белками. Эти белки осуществляют большую часть мембранных функций. Многие из них являются ре-
Глава 1. КЛЕТКА И ТКАНИ

|
|
Фосфолипид
Цепь аминокислоты
Карбогидратная группа
Рис. 1.1.4. Биохимическая организации цитоплазматической мембраны:
/ — карбогидратная группа гликопротеина; 2 — периферический протеин; 3 — карбогидратная группа протеина; 4 — карбогидратная группа гликолипида; 5 — наружная поверхность клетки; 6 — внутренняя поверхность клетки; 7 — трансмембранные белки
цепторами, ферментами, переносчиками различных молекул.
Описанная структура мембраны предопределяет многие важные для жизнедеятельности клетки функции. Вот некоторые из них. Элементарная мембрана обладает избирательной проницаемостью (транспортная функция). При этом вещества могут проходить через нее путем диффузии (молекулы небольшого размера) или при использовании специальных механизмов активного переноса с затратой энергии (крупные полярные молекулы). Диффузия осуществляется по градиенту концентрации, т. е. вещества перемещаются из зоны высокой концентрации в зону низкой концентрации путем броуновского движения.
Активный транспорт осуществляется при помощи белков-переносчиков с использованием энергии АТФ. Происходит он против градиента концентрации вещества.
Функцией цитоплазматической мембраны является также распознание данной клеткой других клеток и прикрепление к ним, взаимодействие с сигнальными молекулами (гормоны медиаторы, цитокины и др.), обеспечение движения клетки благодаря связи плазмолеммы с сократимыми элементами цитоскелета (образование псевдо-, фило- и ламеллоподий).
Описывая цитоплазматическую мембрану, необходимо указать на то, что в настоящее время рассматривают комплекс структур, отделяющих содержимое цитоплазмы от окружающего межклеточного пространства (поверхностный комплекс). Помимо уже упомянутой плазмолеммы, к поверхностному комплексу относят также гликокаликс и премембранные образования цитоскелета.
Гликокаликс располагается на наружной поверхности цитолеммы (рис. 1.1.5). Его толщина колеблется от 8 до 200 нм. Он представляет собой комплекс молекул, связанных с белками
мембраны, и состоит из полисахаридов, глико-липидов и гликопротеинов. Многие из молекул гликокаликса функционируют как специфические молекулярные рецепторы. Именно благодаря рецепторам на поверхности клетки могут закрепляться так называемые сигнальные молекулы, например гормоны.

Рис. 1.1.5. Строение ресничек и поверхностный комплекс эпителиальной клетки:
/ — микроворсинки; 2 — актиновые филаменты в цитоплазме микроворсинок; 3 — плазмолемма микроворсинки; 4 — гликокаликс на поверхности эпителиальной клетки
К внутренней поверхности плазмолеммы примыкают поверхностные структуры цитоплазмы, обеспечивающие передачу информации более глубоко расположенным структурам клетки и запускающие сложные цепи биохимических реакций.
Эндоплазматический ретикулум и рибосомы.Используя электронную микроскопию, в цитоплазме удалось обнаружить гранулярные и трубчатые структуры, формирующие сеть. Эта
Клетка
 сеть была названа эндоплазматическим ретику-лумом (рис. 1.1.6, 1.1.7).
сеть была названа эндоплазматическим ретику-лумом (рис. 1.1.6, 1.1.7).
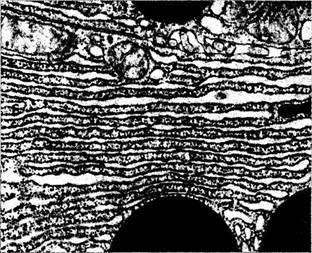
Рис. 1.1.6. Ультраструктурное строение эндоплазматического ретикулума (гранулярного):
видны многочисленные рибосомы, расположенные на мембранах
Система цистерн эндоплазматического ретикулума клетки обладает четко организованной структурой. Цистерны плотно упакованы
рибонуклеопротеидов. В тех случаях, когда на мембранах эндоплазматического ретикулума обнаруживаются многочисленные рибосомы, рети-кулум называют гранулярным (шероховатым) эндоплазматическим ретикулумом. Если рибосом нет, то ретикулум называют агрануляр-ным (гладким) эндоплазматическим ретикулумом.
Рибосомы могут свободно лежать в цитоплазме или формировать маленькие розетки (полисомы). Базофилия цитоплазмы, выявляемая в некоторых типах клеток, связана именно с присутствием рибосом. В некоторых типах нейронов (в частности, ганглиозные клетки сетчатки) отдельным компактным скоплениям цистерн гранулярного эндоплазматического ретикулума на светооптическом уровне соответствуют очерченные участки базофилии цитоплазмы, которые в совокупности называются хромофильной субстанцией, или тельцами Ниссля.
Основной функцией эндоплазматического ретикулума является синтетическая, а именно синтез белков, углеводов, липидов. Для этого в рибосомах существуют все необходимые компоненты: аминокислоты, транспортная РНК и матричная РНК. Смысл тесной связи рибосом с цистернами ретикулума сводится к тому, что при синтезе веществ, подлежащих выведению
|
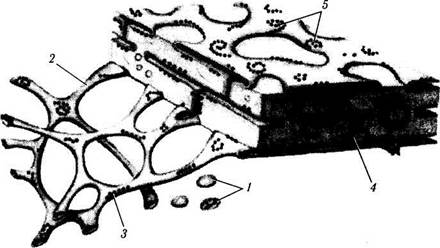
Рис. 1.1.7. Объемная схема организации эндоплазматического ретикулума: / — пузырьки; 2 — тубулярные структуры; 3 — рибосомы; 4 — цистерны; 5 — полисомы
и обычно лежат параллельно друг другу. Они ограничены мембраной. В цистерне виден элек-тронноплотный зернистый материал — матрикс, а иногда и секрет.
На наружной поверхности части мембран располагаются многочисленные маленькие темные частицы диаметром 15 нм, называемые рибосомами. Состоят они преимущественно из
из клетки (например, железы), синтезируемый материал попадает в цистерны, где и окружается мембраной. При этом секрет не попадает в цитоплазму. Участвует эндоплазматический ретикулум также в детоксикации экзогенных и эндогенных веществ, накоплении ионов кальция (в основном, в мышечных клетках), восстановлении кариолеммы в телофазе митоза.
Глава 1. КЛЕТКА И ТКАНИ
 Аппарат (комплекс) Гольджи. Аппарат Гольджи представляет собой вторую мембранную систему клетки, которая не контактирует с эндоплазматическим ретикулумом (рис. 1.1.8).
Аппарат (комплекс) Гольджи. Аппарат Гольджи представляет собой вторую мембранную систему клетки, которая не контактирует с эндоплазматическим ретикулумом (рис. 1.1.8).
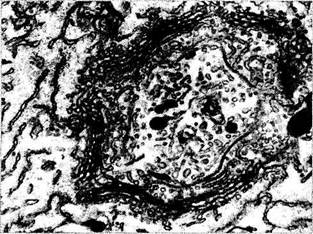
Рис. 1.1.8. Ультраструктурная организация комплекса Гольджи
Чаще всего аппарат Гольджи располагается вблизи ядра и обнаруживается во всех типах клеток. Наиболее развит он в интенсивно сек-ретирующих клетках. В эпителиальных клетках аппарат Гольджи располагается в апикальной части.
Аппарат Гольджи состоит из трех основных компонентов:
1. Стопок уплощенных мешочков (цистерн).
2. Пузырьков.
3. Вакуолей, или секреторных пузырьков
(рис. 1.1.9).
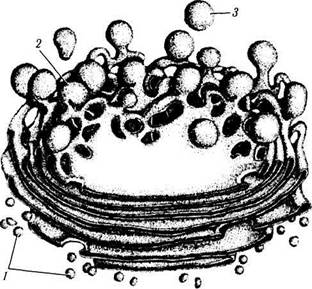
Рис. 1.1.9. Объемная схема комплекса Гольджи (по Leblond):
I — транспортные пузырьки; 2 — зрелая поверхность мешочков; 3 — секреторные пузырьки
В отличие от эндоплазматического ретукулу-ма на мембранах аппарата Гольджи рибосом не выявляется. Образование, состоящее из вышеперечисленных трех структурных элементов, называют диктиосомой (dyctios — сеть). Число диктиосом колеблется в различных клетках от одной до нескольких сотен.
Уплощенные мешочки (цистерны) плотно прилежат друг к другу, образуя как бы стопку (3—30 элементов). Между цистернами определяется пространство, равное 15—30 мкм. Каждая группа цистерн внутри стопки отличается особым составом ферментов. Периферические отделы цистерн несколько расширены и от них отщепляются пузырьки и вакуоли.
Пузырьки, окруженные мембраной, имеют диаметр 40—80 нм и образуются путем отщепления от цистерн. Вакуоли (диаметр — 0,1 — 1,0 мкм) содержат секрет умеренной плотности, находящийся в процессе конденсации.
Та сторона комплекса Гольджи, с которой в него поступают вещества, секретируемые эндоплазматическим ретикулумом, называется цис-полюсом (формирующаяся поверхность), а противоположная — транс-полюсом (зрелая поверхность). Таким образом, аппарат Гольджи структурно и биохимически поляризован.
Основной функцией аппарата Гольджи является его прямое участие в секреторной деятельности клетки (синтез полисахаридов, гликопро-теинов, конденсация секреторного продукта, обеспечение новообразованных гранул мембраной и упаковка в нее секреторных продуктов, сортировка белков на поверхности мембран цистерн и др.).
Функционирование комплекса Гольджи в настоящее время представляется следующим образом. Как указано выше, в гранулярном эндоплазматическом ретикулуме при помощи рибосом осуществляется синтез веществ (рис. 1.1.10). Образовавшийся секрет заключается в мембрану и образует транспортный пузырек, который отделяется от эндоплазматического ретикулума и сливается с цистернами аппарата Гольджи, передавая им свое содержимое. В аппарате Гольджи по мере продвижения от цистерны к цистерне происходит активное химическое преобразование секрета. После преобразования секрета от аппарата опять-таки отделяется пузырек, но уже секреторный, который продвигается к цитоплазматической мембране, сливается с ней, и секрет выделяется наружу. Описанный процесс называется экзо-цитозом.
Судьба пузырьков, отщепляющихся от аппарата Гольджи, различна. Одни из них направляются к поверхности клетки и выводят синтезированные вещества в межклеточный матрикс. Часть этих продуктов является метаболитами, а часть — специально синтезированными веществами, обладающими биологической активностью (секреты).
Клетка

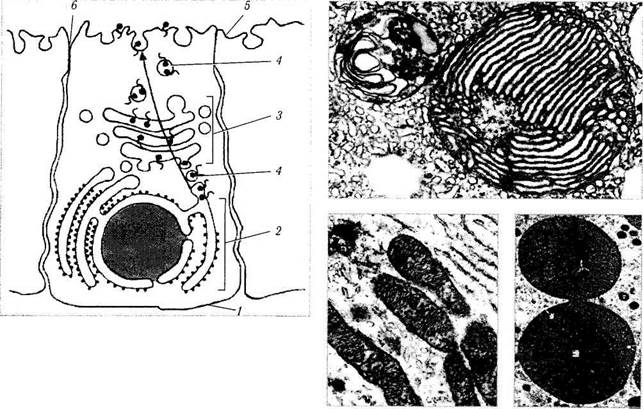 Рис. 1.1.10. Схематическое изображение синтетического аппарата клетки:
Рис. 1.1.10. Схематическое изображение синтетического аппарата клетки:
/ — базальная плазматическая мембрана клетки; 2 — шероховатый эндоплазматический ретикулум; 3 — комплекс Гольджи; 4— транспортные пузырьки; 5 — апикальная плазматическая мембрана; 6 — межклеточное пространство
Аппарат Гольджи участвует в образовании лизосом, важных внутрицитоплазматических органоидов, строение и функции которых будут описаны ниже.
Митохондрии. Помимо эндоплазматическо-го ретикулума и рибосом, в цитоплазме обнаруживаются митохондрии.
При ультраструктурном исследовании митохондрии выглядят удлиненными, сферическими, а иногда ветвистыми образованиями, стенка которых окружена двойной мембраной (рис. 1.1.11). Размер митохондрий самый разнообразный.
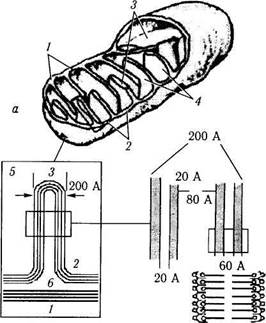
Внутренняя мембрана отдает внутрь органоида многочисленные выпячивания — кристы, благодаря которым площадь внутренней мембраны митохондрий существенно увеличивается. Форма крист в митохондриях большинства клеток пластинчатая (рис. 1.1.12). В некоторых клетках встречаются кристы в виде трубочек и пузырьков (тубулярно-везикулярные кристы).
В пространстве между кристами (митохонд-риальный матрикс) располагаются темные гранулы (30—50 нм в диаметре) — митохондри-альные гранулы (оксисомы или F1-частицы). В этих частицах сосредоточены АТФ-азы — ферменты, непосредственно обеспечивающие распад АТФ. Эти процессы непосредственно связаны с циклом трикарбоновых кислот (цикл Кребса).
В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления
Рис. 1.1.11. Различные морфологические типы митохондрий (электронная микроскопия)
Липид
S В Белок Белок
Рис. 1.1.12. Объемное изображение ультраструктурной организации митохондрии (а) и строение крипты (б, в):
а — схема расположения наружной (/) и внутренней (2) мембран, крист (3) и матрикса (4); б— митохондриальная криста при большом увеличении (5 — внутренняя полость; 6 — наружная полость); в — молекулярная структура кристы
Глава 1.КЛЕТКА И ТКАНИ
 энергии, например вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонем, ресничек), компонентов синтетического аппарата (цистерн эндоплазма-тической сети).
энергии, например вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонем, ресничек), компонентов синтетического аппарата (цистерн эндоплазма-тической сети).
Митохондрии, в отличие от других органелл, обладают собственной генетической системой, необходимой для их самовоспроизведения и синтеза белков. В них обнаруживаются ДНК, РНК и рибосомы. Митохондрии способны размножаться в клетке путем деления. Тем не менее генетическая информация, содержащаяся в ДНК, не обеспечивает их всеми необходимыми белками. Часть этих белков кодируется ядерными генами. Поэтому митохондрии в отношении их самовоспроизведения называют полуавтономными структурами. У человека и других млекопитающих митохондриальный геном наследуется от матери.
Митохондриальная ДНК довольно часто повреждается, что является причиной развития так называемых митохондриальных болезней. Повреждение ДНК происходит в результате образования в матриксе большого количества биоокислителей (перекись водорода, радикалы кислорода). Вследствие этого вероятность мутации митохондриальной ДНК в 10 раз выше ядерной. Мутации митохондриальной ДНК вызывают ряд заболеваний с широким спектром клинических проявлений (слепота, глухота, нарушение движений, сердечная недостаточность, диабет, патология печени и почек и др.). Диагноз некоторых митохондриальных болезней может быть поставлен при изучении биоптата мышечной ткани, в которой выявляются аномальные митохондрии.
Как было указано выше, основной функцией митохондрий является снабжение клетки энергией путем окислительного фосфорилирования, превращая АДФ в АТФ. Более подробные сведения относительно строения и функций митохондрий можно найти в табл. 1.1.2.
Дата добавления: 2015-03-26; просмотров: 873;