АТАКСИЯ, НАРУШЕНИЯ РАВНОВЕСИЯ И ПОХОДКИ
Сид Гилман (Sid Gilman)
Начиная лечение больных с неврологическими расстройствами, необходимо прежде всего установить, имеются ли в анамнезе данные об изменениях поз и походки, а также исследовать эти функции при осмотре. Изменения позы и походки могут возникать в -результате поражения нервной системы на различных уровнях, и часто тип клинических изменений указывает на локализацию поражения.
Образования нервной системы, контролирующие стояние и ходьбу. Образованиями центральной нервной системы, контролирующими положение стоя и походку, являются базальные ядра, «локомоторная зона» среднего мозга, мозжечок и спинной мозг. Важную роль, несомненно, играет кора больших полушарий, однако в экспериментах на животных при полном удалении коры головного мозга в периоде новорожденности с сохранением базальных ядер, зрительного бугра и нижележащих образований стояние и ходьба остаются возможными. Электростимуляция так называемой локомотивной зоны среднего мозга приводит к возникновению движений, характерных для ходьбы, а скорость и манера движений могут изменяться от медленной ходьбы до быстрого шага или бега соответственно изменениям интенсивности раздражения. Эта зона получает проекции из базальных ядер, включая субталамическое и эндопедункулярное ядра и черное вещество.
Нервные образования, координирующие движения мышц, располагаются в спинном мозге. После экспериментальной перерезки спинного мозга на уровне средних грудных сегментов задние конечности, помещенные на движущуюся дорожку, сохраняют способность выполнять координированное передвижение. При увеличении скорости движения дорожки движения видоизменяются и напоминают таковые при беге. При высокой перерезке спинного мозга и верхние, и нижние конечности могут генерировать согласованные чередующиеся движения. Таким образом, нервная система спинного мозга может координировать движения всех четырех конечностей. Многие движения, необходимые для ходьбы, контролируются мозжечком, удаление его приводит к выраженным расстройствам стояния и ходьбы.
Таким образом, ходьба является результатом совместной деятельности базальных ядер, среднего мозга, мозжечка и спинного мозга. Чувствительные сигналы от движений суставов и афферентного аппарата мышц обеспечивают важные составляющие контроля за ходьбой. Без соответствующей чувствительной информации по типу обратной связи походка может значительно изменяться.
Мозжечок. Мозжечок сам по себе не инициирует движений, однако, взаимодействуя с двигательной корой, базальными ядрами и некоторыми стволовыми образованиями, он участвует в контроле за выполнением большинства движений. Мозжечок необходим для поддержания правильного положения тела и устойчивости при ходьбе и беге, для совершения тонких целенаправленных движений, например выполняемых в процессе письма, одевания и приема пищи; для быстрого выполнения чередующихся повторных движений, например игра на музыкальных инструментах или работа на компьютере; для согласования следящих целенаправленных движений глаз. Мозжечок контролирует некоторые качественные показатели движения, такие как направление, скорость и ускорение. Произвольные движения могут существовать и при нарушении деятельности мозжечка, однако они будут неловкими и неорганизованными. Двигательные расстройства, вызываемые нарушением деятельности мозжечка, называют диссинергией (а также аснергией или атаксией).
Мозжечок состоит из центрально расположенного червя и двух полушарий, прилегающих к продолговатому мозгу, мосту и среднему мозгу и соединяющихся с ними с помощью трех ножек с каждой стороны. Поверхность мозжечка покрывает кора мозжечка, или слой серого вещества, окружающий расположенное внутри белое вещество. Внутри мозжечка располагаются три пары глубоких ядер мозжечка, порядок расположения их от центра к периферии следующий: ядро шатра, промежутчатое (пробковидное и шаровидное) и зубчатое ядра.
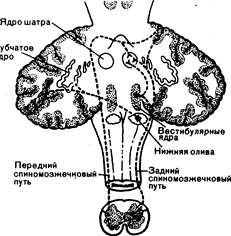
Рис. 16.1. Связи переднего и заднего спиномозжечкового путей с ядрами шатра и вестибулярными ядрами (из S. Gilman, S. S. Winans. — In Manter and Gatz s Essentials of Clinical Neuroanatomy and Neurophysiology, 6th ed. Philadelphia: Davis Co., 1982).
Мозжечок состоит из трех долей. Клочково-узелковая доля, филогенетически являющаяся самой старой частью мозжечка, состоит из парных клочка и узелка. Она получает проекции преимущественно из вестибулярных ядер. Передняя доля, также старая часть мозжечка, состоит из червя и расположенных около него образований в передневерхней части мозжечка. Она получает проекции главным образом из спинного мозга. Задняя доля является самой большой и филогенетически наиболее молодой частью мозжечка и располагается между двумя другими долями. Задняя доля получает проекции из больших полушарий головного мозга через ядра моста.
Кора мозжечка состоит из трех слоев: поверхностного молекулярного слоя, среднего слоя клеток Пуркинье и внутреннего зернистого слоя. Афферентные волокна, достигающие коры мозжечка, посылают коллатеральные проекции в глубокие ядра мозжечка и оканчиваются либо в зернистом слое в виде моховидных волокон, либо на дендритах клеток Пуркинье в виде ползучих волокон. Афферентные сигналы в моховидные волокна приходят от спинного мозга, ядер моста, вестибулярных рецепторов, ядер тройничного нерва, ретикулярной формации и глубоких ядер мозжечка. Афферентная иннервация к ползучим волокнам приходит исключительно от нижних олив. Импульсы, приходящие к моховидным и ползучим волокнам, являются возбуждающими для глубоких ядер мозжечка и коры мозжечка. Клетки Пуркинье обеспечивают единственный путь для всей информации, исходящей из коры мозжечка, и являются возбуждающими для глубоких мозжечковых и вестибулярных ядер.
Мозжечок также получает проекции из голубоватого места и ядер шва ствола головного мозга. Афферентные проекции в мозжечок из голубоватого места являются норадренергическими; из ядер шва — серотонинергическими; и те и другие оказывают тормозное влияние. Предполагаемыми нейротрансмиттерами мозжечка являются некоторые аминокислоты: глутаминовая, используемая зернистыми клетками; аспарагиновая, используемая ползучими волокнами, и гамма-аминомасляная, используемая клетками Пуркинье, Гольджи и корзинчатыми клетками.
Нижняя ножка мозжечка (веревчатое тело) состоит в основном из афферентных волокон. В ножке проходит один эфферентный путь, шатрово-бульбарный, содержащий проекции к вестибулярным ядрам и завершающий вестибулярный круг, проходящий через мозжечок. Афферентные волокна приходят в нижнюю ножку мозжечка по меньшей мере из шести источников (рис. 16.1): волокна из вестибулярного нерва и его ядер; волокна оливомозжечкового пути из нижних оливных ядер; задний спиномозжечковый путь; некоторые из волокон заднего спиномозжечкового пути; клиновидно-мозжечковый пучок из добавочного ядра клиновидного пучка в продолговатом мозге; ретикуломозжечковые волокна. Средняя ножка мозжечка почти полностью состоит из перекрещенных афферентных волокон, идущих от ядер моста в сером веществе основания моста (мостомозжечковые или поперечные волокна моста) (рис. 16.2). В ядра моста ведущие проекции приходят из коры головного мозга. Верхняя ножка мозжечка состоит в основном из эфферентных проекций мозжечка. Из зубчатого и промежуточного ядер идут проекции к красному ядру, зрительному бугру и ретикулярной формации. Часть волокон шатрово-бульбарного пути также на некотором протяжении проходит в составе верхней ножки, до того как достигнет нижней ножки мозжечка. Верхняя ножка мозжечка содержит афферентные проекции переднего спиномозжечкового пути и проекции в мозжечок из ядер тройничного нерва.
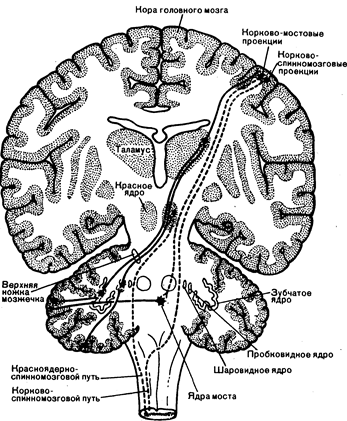
Рис. 16.2. Центральные связи зубчатого и промежуточного (пробковидного и шаровидного) ядер (из S. Cilman, S. S. Winans. — In Manter and Gatz Essentials of Clinical Neuroanatomy and Neurophysiology, 6th ed. Philadelphia: Davis Co., 1982).
За исключением прямых проекций клеток Пуркинье в вестибулярные ядра, эфферентные пути мозжечка начинаются от глубоких ядер. От ядра шатра отходят волокна к вестибулярным ядрам и ядрам ретикулярной формации ствола мозга. Эти ядра проецируются в спинной мозг и имеют отношение к управлению положением тела в пространстве и устойчивостью. Промежуточные ядра с каждой стороны посылают аксоны через верхние ножки мозжечка к красному ядру противоположной стороны. Красное ядро осуществляет связь со спинным мозгом посредством красноядерно-спинномозгового пути (см. рис. 16.2). Этот путь пересс кает среднюю линию и проходит вниз к спинному мозгу. Начало этого пути в промежуточных ядрах и окончание его в спинном мозге находятся с одной стороны. Зубчатое и промежуточное ядра посылают проекции через верхнюю ножку мозжечка к контралатеральному вентролатеральному ядру зрительного бугра. Вентролатеральное ядро дает проекции к двигательным зонам лобной доли на одноименной стороне. Окончания волокон, идущих от зрительного бугра, образуют в коре головного мозга связи с кортико-спинальными нейронами, эфферентные волокна которых проходят в составе пирамидного пути и переходят на противоположную сторону в спинном мозге. Таким образом, начало мозжечково-таламокоркового пути в зубчатом и промежуточном ядрах и его окончание в спинном мозге находятся на одной стороне (см. рис. 16.2).
Для клинических целей принят метод описания мозжечка, основывающийся на наличии продольных сагиттально расположенных зон. Каждая половина мозжечка разделяется на три продольные полоски, расположенные по направлению от центра к периферии, в них входят кора мозжечка, прилежащее белое вещество и глубокие ядра мозжечка. Различают срединную зону, состоящую из области червя и ядра шатра; промежуточную зону, включающую образования, расположенные вокруг червя, и промежуточные ядра; латеральную зону, состоящую из полушария мозжечка и зубчатого ядра. Поражения срединной зоны приводят к изменениям поз и походки, атаксии туловища и неустойчивости при ходьбе, а также к вынужденному повороту или запрокидыванию головы. Повреждения латеральной зоны приводят к нарушениям координированных движений в конечностях (атаксии), дизартрии, гипотонии, нистагму и- кинетическому тремору. Поражения промежуточной зоны вызывают симптоматику, характерную для повреждений обеих (срединной и латеральной) зон.
Атаксия возникает в результате дисметрии и несоразмерности движений. Дисметрией называют нарушение направления или положения конечности во время активного движения, при котором конечность опускается, не достигнув цели (гипометрия), или продвигается дальше цели (гиперметрия). Несоразмерность движений означает ошибки в последовательности и скорости отдельных составляющих движения. В результате возникает потеря скорости и ловкости движений, требующих плавной совместной деятельности различных мышц. Движения, бывшие до того плавными и точными, становятся неровными и неточными. Клинически атаксия бывает представлена в виде нарушений темпа и объема отдельных движений и обычно возникает при поражении мозжечка или нарушении различных видов чувствительности. Атаксия ходьбы характеризуется неравномерным темпом, продолжительностью и последовательностью движений с пошатыванием из стороны в сторону.
Физиологические реакции, имеющие значение для стояния и ходьбы. Удерживание туловища в вертикальном положении осуществляется посредством многочисленных механизмов, реагирующих на изменение постуральных рефлексов: локальными статическими реакциями отдельных конечностей; сегментарными статическими реакциями, обеспечивающими согласованность движений; общими статическими реакциями, возникающими при перемещении головы в пространстве. К локальным статическим реакциям относятся реакции растяжения мышцы и положительные реакции удержания положения. Простейший рефлекс растяжения мышцы можно представить в виде напряжения мышцы (глубокий сухожильный рефлекс), кратковременного мышечного сокращения, вызванного резким натяжением сухожилия мышцы. Удержание мышцы в состоянии растяжения приводит к длительному сокращению этой мышцы, происходящему благодаря рефлексу растяжения. Положительная реакция удержания положения, как было установлено в экспериментах на животных, возникает вследствие контакта кожи ступни, а также натяжения межкостных мышц, приводящего к возникновению проприоцептивной стимуляции. В результате этого раздражения возникает разгибательный толчок в конечности.
К сегментарным статическим реакциям относят перекрестный разгибательный рефлекс и согласованность движений конечностей. При перекрестном разгибательном рефлексе чрезмерное раздражение конечности приводит к ее сгибанию и одновременному разгибанию противоположной конечности. При более интенсивном раздражении перекрестный разгибательный рефлекс, приводящийся в действие пораженной задней конечностью, может вызвать сгибание противоположной передней конечности и разгибание гомолатеральной передней конечности. Таким образом, все туловище движется по диагонали благодаря разгибанию контралатеральной задней конечности и гомолатеральной передней конечности, удаляя раздражаемую конечность от источника раздражения. Данный диагональный характер согласованных движений обеспечивает изменение положения тела в различных ситуациях.
Общие статические рефлекторные реакции делятся на два типа. В первом случае тонические шейные и лабиринтные рефлексы совместно обеспечивают регуляцию положения тела при различных движениях головы относительно туловища, во втором установочный рефлекс возникает при лабиринтных, шейных и зрительных раздражениях и помогает животному восстановить вертикальное положение после падения. Одним из компонентов установочного рефлекса является хватательный рефлекс. Среди других видов общих статических реакций отмечают рефлексы положения и прыгания, а также регулирование положения тела во время движения конечностей.
Дата добавления: 2015-03-17; просмотров: 1455;