БИОСИНТЕЗ НУКЛЕОТИДОВ
Существует 2 пути биосинтеза нуклеотидов в клетке. Во-первых, путь повторного использования АО и нуклеозидов (не только экзогенных, но и образовавшихся в клетке в процессе репарации ДНК или при распаде «отработавших» РНК). Наиболее активно протекает в клетках интенсивно размножающихся тканей (эмбриональных, регенерирующих, эпителиальных, опухолевых). Во-вторых, синтез de novo (из низкомолекулярных предшественников).
Пути повторного использования АО и нуклеозидов: наличие этих путей позволяет использовать синтетические аналоги пуринов и пиримидинов для химиотерапии опухолей и лечения вирусных инфекций (например, 5-фторурацил, меркаптопурин, ацикловир, АЗТ и др.). Такие препараты включаются клеткой в состав нуклеотидов, встраиваются в молекулу ДНК и вызывают цитотоксический эффект.
| |||
 |
 1. Нуклеозид Нуклеозидмонофосфат НТФ.
1. Нуклеозид Нуклеозидмонофосфат НТФ.
Нуклеозидкиназа
Чаще используется для повторного использования пиримидинов (тимидинкиназа, цитидинкиназа).
2. Синтез нуклеотидов на основе готовых азотистых оснований больше характерен для пуринов и проходит в 2 этапа:
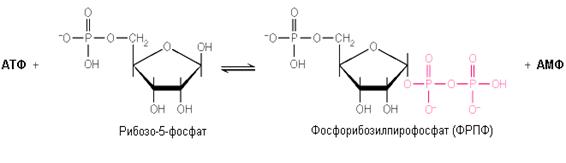 |
а) образование активной формы рибозо-5-фосфата (фосфорибозилпирофосфата):
б) взаимодействие ФРПФ с азотистым основанием:
 АО
АО  Нуклеозидмонофосфат НТФ
Нуклеозидмонофосфат НТФ
Фосфорибозил
трансфераза
De novoсинтез пуриновых нуклеотидов
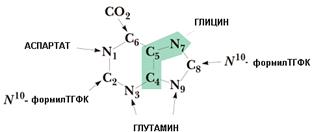 Особенностью синтеза пуринов de novo является то, что за основу берется рибозо-5-фосфат и на его основе формируется пуриновое кольцо. N-гликозидная связь формируется уже на ранних этапах синтеза.
Особенностью синтеза пуринов de novo является то, что за основу берется рибозо-5-фосфат и на его основе формируется пуриновое кольцо. N-гликозидная связь формируется уже на ранних этапах синтеза.
Источником всех атомов азота для пуринового ядра являются аминокислоты (глицин, глутамин, аспартат). Источники атомов углерода: СО2 и формил-ТГФК (активная форма фолиевой кислоты, В9). Общим предшественником для адениловых и гуаниловых нуклеотидов является инозинмонофосфат (ИМФ).
 Рибозо-5-фосфат + АТФ Фосфорибозилпирофосфат + АМФ
Рибозо-5-фосфат + АТФ Фосфорибозилпирофосфат + АМФ
 ФРПФ- (ФРПФ)
ФРПФ- (ФРПФ)
синтетаза
Амидотрансфераза
Фосфорибозиламин
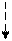
+гли
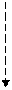 +глн + СО2
+глн + СО2
+асп + 2формил-ТГФК
Инозинмонофосфат
 | |||
 |
+асп+глн
+ГТФ+АТФ
 АМФ ГМФ
АМФ ГМФ

АДФ ГДФ
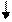
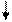
АТФ ГТФ
Ключевой фермент синтеза пуринов: амидотрансфераза.
Регуляция:
1) аллостерическая: избыток конечных продуктов (АТФ, ГТФ) ингибирует ключевой фермент; избыток пиримидиновых нуклеотидов его активирует;
2) ГМФ ингибирует образование ксантиловой кислоты, а АМФ — аденилоянтарной;
3) перекрестная: для синтеза АМФ требуется ГТФ, а для синтеза ГМФ требуется АТФ;
Наиболее распространенной формой нарушения обмена пуринов является подагра. Основная причина — повышение уровня мочевой кислоты в крови (гиперурикемия) и ее отложение в почках и суставах. Причины: а) избыточный синтез пуриновых нуклеотидов (нечувствительность ферментов к регуляторам); б) дефект ферментов реутилизации пуринов;
в) патология почек (недостаточное выведение). Способствует избыточное потребление пуринов с пищей. В лечении подагры используется аллопуринол — ингибитор ксантиноксидазы.
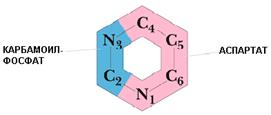 De novo синтез пиримидиновых нуклеотидов
De novo синтез пиримидиновых нуклеотидов
В отличие от пуринов, при биосинтезе пиримидинов de novo вначале образуется пиримидиновое кольцо, а лишь затем к нему присоединяется рибозо-5-фосфат. Источниками атомов для пиримидинового кольца являются глутамин, аспартат и СО2. Синтез начинается с образования карбамоилфосфата:
Глутамин + СО2 + 2 АТФ ¾¾¾¾¾¾® Карбамоилфосфат + Глутамат + 2 АДФ + Фн.
Карбамоилфосфат-
синтетаза II
В отличие от карбамоилфосфатсинтетазы I, фермент синтеза пиримидинов использует амидный азот глутамина (а не свободный аммиак) и локализован в цитоплазме.
 Карбамоилфосфат + Аспартат ¾¾¾¾¾¾® Карбамоиласпарагиновая Аспартаткарбамоил- кислота
Карбамоилфосфат + Аспартат ¾¾¾¾¾¾® Карбамоиласпарагиновая Аспартаткарбамоил- кислота
трансфераза
Оротовая к-та
 +ФРПФ
+ФРПФ
Оротидин-5/-мономофосфат

 ДГФК метилен-ТГФК –СО2
ДГФК метилен-ТГФК –СО2
Дата добавления: 2015-03-11; просмотров: 2994;