ПРОТИВОИНФЕКЦИОННАЯ ЗАЩИТА РЕПРОДУКТИВНЫХ ОРГАНОВ ЖЕНЩИНЫ
Урогенитальная инфекция (УГИ) является основной причиной возникновения многих заболеваний, встречающихся в акушерстве и гинекологии. Отрицательное действие ее многогранно. УГИ вызывает не только воспалительные заболевания женских половых органов, что само по себе приводит к различным тяжелым осложнениям, но и является фактором патогенеза других невоспалительных заболеваний: миомы, эндометриоза, опухолей яичников и др. Кроме того, УГИ может приводить к аутоиммунным процессам в организме с развитием ДВС-синдрома. Во время беременности УГИ обусловливает возникновение многих осложнений, среди которых особое значение имеют невынашивание беременности и внутриутробное инфицирование плода.
В настоящее время большое внимание уделяется противоинфекционной резистентности организма, в которой условно выделяют системные и локальные факторы (Кузник Б.И., Васильев Н.В., Цибиков Н.Н., 1989; Нуруше-ваС.М., 1996; Пауков B.C., ГостищевВ.К. и др., 1996).
Исследование особенностей местного иммунитета у больных с ВЗОМТ получило широкое распространение только в последние годы, в связи с чем в
Генитальные инфекции и факторы противоинфекционной защиты в генезе
воспалительных заболеваний женских половых органов 25
литературе данные о показателях местного иммунитета слизистых оболочек полового тракта немногочисленены (Митков В.Г., 2004).
По мнению многих авторов (Говалло В.И., 1987; Теплякова М.В., Радион-ченкоА.А., Рыжова И.А., 1990; Медведев Б.И., Долгушина В. Ф., 1993;Хмель-ницкий O.K., 1994; Долгушина В.Ф., Телешева Л.Ф., Смольникова Л.А., 1995; ДергачеваТ.И., 1996; Долгушина В.Ф., Телешева Л.Ф. и др., 1996; Сапин М.Р., Этинген Л.Е., 1996; Долгушин И.И., Долгушина В.Ф., Телешева Л.Ф., 1997), антимикробная устойчивость генитального тракта обеспечивается сложным защитным комплексом, в состав которого входят:
• анатомо-физиологические особенности строения наружных половых органов;
• нормальная микрофлора влагалища, которая обеспечивает колонизационную резистентность генитального тракта;
• микроэкология шейки матки, слизистая оболочка которой обладает чрезвычайно высокими бактерицидными свойствами, имеет автономную иммунную систему и является последним барьером для восходящей инфекции;
• состояние реактивности организма на локальном и общем уровнях, к которым относят неспецифические и иммунные факторы защиты.
Неспецифические факторы защиты обеспечивают естественную резистентность организма к инфекциям. Среди них выделяют:
• естественные барьеры — кожа, слизистые оболочки, которые первыми вступают в контакт с возбудителями инфекций. Слизистые оболочки обладают выраженными бактерицидными свойствами, обусловленными продукцией секреторного иммуноглобулина A (slgA) и лизоцима;
• систему фагоцитоза, включающую нейтрофилы и макрофаги. Фагоцитарная функция нейтрофилов оценивается по трем показателям: активности фагоцитоза (процент фагоцитирующих нейтрофилов); интенсивности фагоцитоза (среднее число частиц, поглощенных одним нейтрофилом); завершенности фагоцитоза (индекс переваривания) (Макаров К.В., 1998);
• систему комплемента, представляющую собой комплекс сывороточных белков, который активирует фагоцитоз, участвует в регуляции гуморального иммунитета и синтеза макрофагов;
• бактерицидные гуморальные факторы, к которым относятся лизоцим, фибронектин, лактоферрин и др.;
• систему интерферонов (ИФН), которые значительно опережают ответ иммунной системы, обладают антивирусной, иммуномодулирующей, противоопухолевой активностью;
• систему нормальных киллеров, функционирующих как эффекторы противовирусной защиты.
Специфическая резистентность организма обеспечивается иммунной защитой, основной функции которой является способность отличать собственные структуры от генетически чужеродных (антигенов), перерабатывать и элиминировать их (Долгушин И.И., Долгушина В.Ф., Телешева Л.Ф., 1997; Кетлинский С.А., Калинина И.М., 1998).
Существует 2 типа иммунного ответа: гуморальный и клеточный. В гуморальном ответе основную роль играют В-лимфоциты, которые продуцируют иммуноглобулины, направленные против антигенов; в клеточном — Т-лимфо-циты, которые в ответ на антиген дифференцируются либо в цитотоксические Т-лимфоциты (CD8), осуществляющие киллерную функцию, либо в Т-хелпе-ры (CD4), активирующие цитотоксичность макрофагов, а также принимаю-
 26 Слава I
26 Слава I
щие активное участие в гуморальном ответе. Центральная роль в регуляции иммунных реакций как на локальном уровне слизистых оболочек, так и на уровне всего организма в целом отводится цитокинам, которые являются медиаторами межклеточных взаимоотношений (Долгушина В.Ф., 1991).
Благодаря слаженному функционированию всех факторов резистентности организм надежно защищен от воздействия инфекционных агентов. Поэтому перспективным в своевременной профилактике и лечении воспалительных заболеваний гениталий является изучение факторов противоинфекционной защиты, как на локальном, так и на органном уровнях.
1.3.1. Нормальный микробный биоценоз влагалища
В настоящее время большое значение в развитии воспалительных заболеваний половых органов отводится нормальной микрофлоре влагалища, которая обеспечивает колонизационную резистентность (КР) генитального тракта. КР подразумевает совокупность механизмов, обеспечивающих стабильность количественного и видового состава нормальной микрофлоры, что предотвращает заселение влагалища патогенными микроорганизмами или чрезмерное размножение условно-патогенных микроорганизмов (УПМ), входящих в состав нормального микроценоза, и распространение их за пределы экологической ниши.
В разные периоды жизни женщины в зависимости от активности репродуктивной функции микроценоз влагалища имеет определенные особенности, однако в любом периоде взаимодействие микрофлоры с иммунной системой организма обеспечивает состояние КР.
Вагинальная микроэкосистема состоит из постоянно обитающих (индиген-ная) и транзиторных (случайная микрофлора) микроорганизмов.
Индигенная микрофлора доминирует по численности популяции, хотя количество представляющих ее видов невелико, в отличие от видового разнообразия транзиторных микроорганизмов, общая численность которых в норме не превышает 3—5% от всего пула, составляющего микроценоз. Состояние КР влагалища связывают именно с индигенной микрофлорой, которая у женщин репродуктивного возраста представлена обширной группой лактобацилл. Благодаря специфической адгезии на эпителиальных клетках влагалища образуется биопленка, состоящая из микроколоний лактобацилл, окруженных продуктами их метаболизма — гликокаликсом.
В репродуктивном возрасте состав вагинального отделяемого многокомпонентен, определяется присутствием секрета бартолиновых и парауретральных желез, транссудата стенок влагалища, отторгшихся эпителиальных клеток, слизи цервикального канала, эндометриальной и трубной жидкости. Концентрация различных компонентов вагинальной среды колеблется в зависимости от дня менструального цикла, однако в ней постоянно присутствуют хлориды натрия и калия, сульфаты, витамины, ионы металлов, муцин, ферменты (амилаза, глюко-зидаза, антитрипсин), белки (альбумины и глобулины), комплексные углеводы, липиды, жирные кислоты. Характерны для вагинальной среды также низкое содержание растворимого кислорода (который является продуктом жизнедеятельности микроорганизмов) и низкий редокс-потенциал.
У женщин детородного возраста слизистая оболочка влагалища состоит из 4 слоев эпителиальных клеток: базального, парабазального (митотически активного), промежуточного (гликогенсодержащего) и поверхностного (слу-щивающегося).
Эстроген, вырабатываемый яичниками, стимулирует митотическую активность, созревание эпителиальных клеток и синтез в них гликогена. Эстроген-
Генитальные инфекции и факторы противоинфекционной защиты в генезе
воспалительных заболеваний женских половых органов 27
ные рецепторы располагаются в базальном и парабазальном слоях слизистой оболочки и практически отсутствуют в поверхностных слоях. Количество этих рецепторов колеблется на протяжении менструального цикла. Их больше в про-яиферативной, чем в секреторной фазе цикла, а в детском возрасте и в постменопаузе их количество минимальное. Таким образом, только на фоне достаточного уровня эстрогенов проявляется митотическая активность парабазального слоя и, следовательно, созревание, пролиферация вагинального эпителия. Тем самым эстрогены стимулируют синтез гликогена в промежуточных клетках. В процессе деструкции созревших поверхностных клеток освобождается гликоген, который служит субстратом для бактериальных ферментов. Происходит расщепление гликогена с образованием глюкозы — основного источника энергии для вагинальных лактобацилл. Бактерии утилизируют глюкозу в процессе анаэробной ферментации, конечным продуктом которой является молочная кислота. Именно такой цикл химических превращений определяет кислую реакцию вагинальной среды (рН 3,8—4,5), что благоприятствует выживанию ацидофильных микроорганизмов, в первую очередь — лактобацилл.
Взаимодействие лактобацилл с другими членами вагинального микроценоза носит сложный характер, патогенетические механизмы которого остаются неизученными. Однако бытующее со времен Додерлейна представление о вагинальной микрофлоре как постоянной и однородной, состоящей из лактобацилл, в настоящее время значительно изменилось.
Результаты современных исследований позволяют охарактеризовать вагинальную микроэкосистему как весьма динамичную и многокомпонентную по видовому составу. Строгая эстрогенная зависимость является ее отличительной особенностью и объясняет динамическую изменчивость в разные периоды жизни женщины (отсутствие функциональной активности яичников в детском возрасте, половое созревание, детородный возраст, постменопауза) и на протяжении менструального цикла в репродуктивном возрасте.
Расшифровка, понимание механизмов, обеспечивающих физиологическую роль индигенной микрофлоры влагалища, открывают пути к профилактике различных патологических состояний.
1.3,1.1. Микрофлора влагалища у девочек
У девочек сразу после рождения слизистая оболочка влагалища имеет хорошо выраженные четыре слоя эпителиальных клеток, что связано с высоким уровнем в крови плода половых гормонов, в том числе эстрогенов. Такая эстроге-новая насыщенность носит пассивный характер и объясняется трансплацентарным переходом в кровоток плода половых гормонов, синтезируемых материнским организмом. Эстрогены стимулируют у плода и новорожденной девочки синтез гликогена в промежуточных клетках влагалищного эпителия, что создает благоприятные условия для приживления лактобацилл. Они доминируют в содержимом влагалища в первые недели жизни. Величина рН отделяемого влагалища составляет около 4,0. По мере снижения в крови ребенка концентрации материнских эстрогенов (в течение 4—6 нед.) прекращается эстрогензависимое созревание влагалищного эпителия. Он становится тонким, пролиферации пара-базальных клеток не происходит, синтез гликогена прекращается, исчезают условия для жизнедеятельности лактобацилл. Осуществляется постепенное вытеснение лактобацилл микроорганизмами соседних локусов — кожи промежности и наружных половых органов, прямой кишки. В этот период жизни девочки вплоть до наступления менархе и становления менструальной функции на фоне нефунк-ционирующих яичников в составе микрофлоры влагалища доминируют грампо-



 28 Глава I
28 Глава I
ложительные кокки - эпидермальный и другие коагулазоотрицательные стафилококки, микрококки, негемолитический стрептококк. Реже встречаются непатогенные нейссерии и коринебактерии, еще реже - эшерихии и энтерококки. Показатель рН вагинального содержимого увеличивается до 6,0—6,5.
Исследования показали, что существует тесная взаимосвязь между уровнем физического и полового развития, биоценозом влагалища, становлением иммунной системы (Афанасьев С.С., Алешкин В.А. и др., 2000; Кулаков В.И., Алешкин В.А. и др., 2000; Салолина И.В., 2000; Бойко А.В., Алешкин В.А. и др., 2001).
Микробная флора влагалища у девочек до полового созревания преимущественно кокковая, по мере полового созревания увеличивается количество лак-тобацилл, и у половозрелых девочек микрофлора практически полностью представлена лактобациллами.
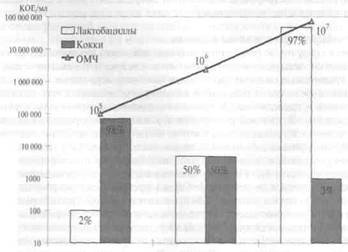
Количественное определение микроорганизмов в вагинальном секрете показало, что у здоровых девочек 6—8 лет общее микробное число (ОМЧ) колеблется от 102 до 105 КОЕ/мл, флора преимущественно кокковая (рис. 1.1). По мере взросления девочки ОМЧ увеличивается и у здоровых девочек старше 9 лет (но не имеющих менструаций) составляет 105—106 КОЕ/мл, лактобацил-лы присутствуют в количестве 102—103 КОЕ/мл. У девочек, менархе у которых уже наступило, микрофлора влагалища становится обильнее, ОМЧ составляет 105—107 КОЕ/мл, в основном это лактобациллы, количество сопутствующих им микроорганизмов обычно не превышает 103—104 КОЕ/мл. Таким образом, изменение микробного числа по мере взросления девочки происходит за счет увеличения количества лактобацилл и некоторого снижения численности кокковой флоры (Бойко А.В., Алешкин В.А. и др., 2001).
|
| 6-8 лет От 9 лет до менархе От менархе до 17 лет Рис. 1.1. Микробиоценоз влагалища у девочек. |
1.3.1.2. Микрофлора влагалища у женщин репродуктивного возраста Изучению влагалищной микрофлоры у женщин репродуктивного возраста посвящено много исследований, начиная с классических работ Додер-
Генитальные инфекции и факторы противоинфекционной защиты в генезе
воспалительных заболеваний женских половых органов 29
лейна (1892). По мере совершенствования микробиологических технологий и проведения комплексных исследований — биохимических, гормональных с учетом клинических, микробиологических и эпидемиологических данных — углублялись представления о микроэкологии вагинального биотопа. Большинство опубликованных работ (в основном ранние публикации) касаются выделения и идентификации бактериальных видов без определения их численности. В меньшем числе работ (более поздние исследования) изучали общую численность всех микроорганизмов, но не отдельных видов. И только в последние годы появились работы, в которых проводят как изучение видового состава, так и определение численности каждого фенотипа, входящего в микроценоз, т.е. имеет место качественная и количественная оценка состава микроценоза.
У здоровых женщин детородного возраста общее количество микроорганизмов в вагинальном отделяемом составляет 6—8,5 lg КОЕ/мл (или на 1 г) и состоит из разнообразных видов, число которых может достигать 40 и более (Муравьева В.В., 1997; Коршунов В.М., Володин Н.Н. и др., 1999; Onder-donk A.B., Wissemann K.W., 1993). Доминирующим родом влагалищной среды являются Lactobacillus spp. (95—98%) — большая группа бактерий, в основном микроаэрофилов, в сочетании с облигатно-анаэробными видами. Несмотря на разнообразие видового состава лактобацилл, выделяемых из влагалища здоровых женщин (более 10 видов), не удается определить ни одного вида, который присутствовал бы у всех женщин (Коршунов В.М., Володин Н.Н. и др., 1999; Onderdonk A.B., Wissemann K.W., 1993). Чаще всего удается выделить следующие виды лактобацилл: L. acidophilus, L. brevis, L.jensenii, L. casei, L. leishmanii, L. plantarum.
Эстрогензависимая способность лактобацилл к адгезии на эпителиальных клетках влагалища, продукция перекиси водорода и антибиотикоподобных веществ, способность при ферментативном расщеплении гликогена образовывать молочную кислоту, снижающую уровень рН до 4,0—4,5, обеспечивают наиболее благоприятные условия для жизнедеятельности лактобацилл и препятствуют размножению ацидофобных бактерий.
Среди транзиторных микроорганизмов влагалища чаще других удается выделить коагулазоотрицательные стафилококки, в первую очередь S. epidermi-dis, Corynebacterium spp., Streptococcus spp., Bacteroides — Prevotella spp., Mycoplasma hominis, которые обычно присутствуют в умеренном количестве (до 4 lg КОЕ/г). Столь же часто, но в меньшем количестве встречаются Micrococcus spp., Propionibacterium spp., Veillonella spp., Eubacterium spp.
Среди сравнительно редко встречающихся микроорганизмов (менее чем у 10% обследованных) обнаруживаются Clostridium spp., Bifidobacterium spp., Actinomyces spp., Fusobacterium spp., Ureaplasma urealyticum, Staphylococcus aureus, Neisseria spp., E. coli и другие колиформные бактерии, Mycoplasma fermentans, Gardnerella vaginalis, Candida spp.
Как видно из этого перечня, видовое разнообразие вагинальной микрофлоры велико, а возможные сочетания микроорганизмов столь многочисленны, что можно говорить лишь о некоторых общих тенденциях. Так, большинство исследователей считают, что у здоровых женщин, кроме лактобацилл, чаще всего присутствуют непатогенные коринебактерии и коагулазоотрицательные стафилококки - у 60-80% обследованных (Соловьева И.В., 1987; Муравьева В.В., 1997; Duerden B.L., 1980). Среди облигатно анаэробных видов большинство исследователей обращает внимание на группу Bacteroides—Prevotella, что связано с высокой их значимостью как этиологических агентов при воспалительных заболеваниях гениталий. Эти микроорганизмы (преимущественно
 30 Глава I
30 Глава I
вид В. bivius) встречаются в низких титрах у 55% здоровых женщин (Masfa-ri A.N., Duerden B.L., 1986).
Микроценоз влагалища является динамичной экосистемой, чутко реагирующей на изменения уровня половых гормонов в крови. Проведенные количественные и качественные исследования вагинальной микрофлоры (Sautter R.L., Brown WJ., 1980; Larsen В., Galask R.P., 1982; Johnson S.R., Petzold C.R., Galask R.P., 1985) также показали, что концентрация и видовой состав микроорганизмов, выделенных в разные фазы менструального цикла, колеблются в значительных пределах. A.B.Onderdonk и соавт. установили, что снижение общего числа бактерий во время менструации происходит в основном за счет уменьшения количества лактобацилл. Численность же облигатных анаэробов из группы бактероидов относительно увеличивается по сравнению с их числом на 21-е сутки менструального цикла. Другие авторы (Sautter R.L., Brown W.J., 1980; Larsen В., Galask R.P., 1982; Johnson S.R., Petzold C.R., Galask R.R, 1985) также отмечают, что во время менструации численность лактобацилл снижается, и они вытесняются разнообразными грамположительными факультативно анаэробными микроорганизмами, такими как коагулазоотрицательные стафилококки, коринебактерии, стрептококки. A.B.Onderdonk и соавт. (1986) проведено уникальное микробиологическое исследование вагинального отделяемого у 8 женщин в течение 9 мес. Среди идентифицированных микроорганизмов были представители 61 вида грамположительных и грамотрицательных бактерий. Авторы обращают внимание на то, что во влагалищной среде может быть обнаружено множество разнообразных фенотипов, но у одной и той же женщины набор этих видов относительно постоянен на протяжении длительного времени. Авторы исследовали 805 проб вагинального отделяемого и идентифицировали более 15 000 микробных культур. Все выделенные бактерии они объединили в 9 основных групп: стафилококки, лактобациллы, бактероиды, пептококки, коринебактерии, стрептококки, эшерихии, анаэробные стрептококки, гардне-реллы. Частота их выделения была различной в разные фазы менструального цикла. При обследовании на 2-й, 4-й и 21-й дни цикла стафилококки выделены из 95,98; 98,75 и 96,17% проб соответственно, а их численность, выраженная в lg КОЕ/г отделяемого, составила 6,72; 7,17 и 5,56. Сходные соотношения получены для группы бактероидов: 89,93; 88,17 и 73,63% в пробах тех же дней цикла с численностью 5,88; 6,02 и 4,0. На 3-м месте по частоте обнаружения в эти дни цикла были лактобациллы — у 78,49; 83,56 и 90,92% обследованных, с численностью популяций соответственно 6,21; 7,08 и 7,91 lg КОЕ/г. Частота выделения микроорганизмов остальных 6 групп (пептококки, коринебактерии, стрептококки, анаэробные стрептококки, эшерихии, гарднереллы) намного ниже: количество этих микроорганизмов составляло от 1,06 до 4,43 lg КОЕ/г.
Суммируя результаты исследований, посвященных связям количественного и видового составов микрофлоры влагалища в зависимости от фазы менструального цикла, можно заключить, что в первые дни менструального цикла увеличивается рН содержимого влагалища до 5,0—6,0. Это связано с появлением большого числа дегенерированных клеток эндометрия и элементов крови. На этом фоне уменьшается обшее количество лактобацилл и относительно увеличивается численность факультативно и облигатно анаэробных бактерий. После окончания менструации популяция лактобацилл быстро восстанавливается и достигает максимального количества в середине секреторной фазы, когда содержание гликогена в эпителии влагалища наиболее велико. Этот процесс сопровождается увеличением содержания молочной кислоты и снижением рН до 3,8—4,5. Во второй фазе менструального цикла абсолютно доминиру-
Генитальные инфекции и факторы противоинфекционной защиты в генезе
воспалительных заболеваний женских половых органов 31
ют лактобациллы, а количество облигатных анаэробов и колиформных бактерий снижается. Приведенные данные дают основание полагать, что в первой (пролиферативной) фазе менструального цикла восприимчивость организма-хозяина к инфекции может возрастать. Этот вывод подтверждается клиническими наблюдениями. Так, по данным H.Thadepalli и соавт. (1982), частота воспалительных осложнений после гистерэктомии у женщин, оперированных в первой фазе менструального цикла, составила 31,6%, а у оперированных во второй фазе - 18%.
1.3.1.3. Микрофлора влагалища во время беременности,
в родах и в послеродовом периоде
Во время беременности под влиянием гормонов желтого тела слизистая оболочка влагалища значительно утолщается. Эластичность клеток промежуточного слоя увеличивается, синтез гликогена в них осуществляется с максимальной интенсивностью. Создаются благоприятные условия для жизнедеятельности лактобацилл. По мере прогрессирования беременности снижается численность транзиторных микроорганизмов и увеличивается количество лактобацилл. В связи с постоянно низкими показателями рН (3,8—4,2) создаются благоприятные условия для количественного увеличения таких микроорганизмов транзиторной группы, как генитальные микоплазмы и дрожжевые грибы. Частота их выделения во время беременности в различных группах риска возрастает до 20—30% и более (Анкирская А.С., Муравьева В.В., Акопян Т.Э., 1994). В литературе последних лет имеются единичные работы, в которых изучали в динамике прогрессирования беременности состояние вагинальной микроэкологии.
При изучении состояния микроценоза цервикального канала в динамике беременности C.P.Goplerud и соавт. (1976) отметили видовое разнообразие выделенных микроорганизмов — 14 факультативных анаэробных видов и 24 облигатных анаэробных. Почти во всех случаях выделялись ассоциации 3-5 видов. Установлена очень важная закономерность с точки зрения защиты организма от инфекции: к сроку родов резко снижалось количество аэробных видов, таких как колиформные бактерии, и таких облигатных анаэробов, как бактероиды и пептострептококки, и нарастало число лактобацилл.
При сроке беременности 8—10 нед. (I триместр) преобладающими микроорганизмами влагалища у женщин с физиологически протекающей беременностью являются лактобациллы, которые выделяются у 96,6% обследуемых в количестве 7,1+0,1 КОЕ/мл. Представители УПМ выделяются с разной частотой, но в значительно меньших количествах. Так, энтеробактерии (во всех случаях типичная кишечная палочка) выделяются лишь у 20,0% беременных в концентрации 3,8+0,1 КОЕ/мл содержимого влагалища. Частота обнаружения стрептококков достигает 53,5%, а их концентрация не превышает 4,7+0,3 КОЕ/мл, облигатно анаэробные бактерии выделяются у 83,3%, но их популяционный Уровень не превышает значений 4,3±0,3 КОЕ/мл.
С увеличением срока беременности (II триместр) в составе вагинальной микрофлоры определяются существенные изменения, которые характеризуются достоверным снижением энтеробактерии до 2,8+0,2 КОЕ/мл (р<0,05) и увеличением количества молочно-кислой микрофлоры в 8,5 раза (р<0,05). Качественный и количественный состав других представителей индигенной вагинальной микрофлоры существенным образом не меняется. Наиболее значительные позитивные сдвиги в составе микробиоценоза влагалища при физиологическом течении беременности отмечаются к 37—38-й неделе бере-
32 Глава I
менности (III триместр). К этому периоду происходит дальнейшее достоверное снижение уровня УПМ - среднее количество энтеробактерий снижается в 80 раз относительно их содержания в I триместре. Частота высеваемости и количественное содержание грибов рода Candida остается к концу беременности на достаточно высоком уровне - 4,5+0,5 КОЕ/мл. Лактобациллы занимают доминирующее положение в микробиоценозе влагалища к концу III триместра беременности, достигая значений 7,9±0,3 КОЕ/мл вагинального содержимого (Кафарская Л.И., 2002).
Таким образом, к сроку родов снижается уровень микробного обсеменения родовых путей при максимальном доминировании лактобацилл, и ребенок рождается в условиях преобладания ацидофильных бактерий, обеспечивающих КР родового канала. В родах плод впервые сталкивается с материнской микрофлорой, которая колонизирует кожные покровы и слизистые оболочки рождающегося ребенка.
Восприятие новорожденным микроорганизмов как «своих» связано с формированием у плода иммунологической толерантности к нормальной микрофлоре. Б.А.Шендеров (1998) на основе анализа зарубежных работ и собственных экспериментальных данных сформулировал гипотезу о формировании иммунологической толерантности к нормальной микрофлоре. Согласно этой гипотезе иммунологическая память к нормальной микрофлоре формируется внутриутробно и полностью зависит от состава микрофлоры матери. Бактериальные антигены, освобождающиеся из микробных клеток в результате физиологического метаболизма или различных воспалительных процессов, проникают в кровяное русло беременной женщины и через плацентарный барьер попадают в кровоток плода. Достигая тимуса плода, антигены вызывают специфическую стимуляцию предшественников Т-супрессоров, которые могут распознавать эти антигены. В каком направлении после рождения пойдет развитие иммунного ответа — по типу иммунологической толерантности (будет происходить приживление нормальной микрофлоры) или иммунного ответа с синтезом IgA (реакция на чужеродные антигены), — зависит от количества возникших в тимусе клеток-предшественников и их специфичности. А это в свою очередь определяется массивностью антигенной дозы, достигшей тимуса плода, степенью общности микробных и тканевых антигенов, сроком беременности (т.е. длительностью микробного воздействия на клетки тимуса). В процессе внутриутробного развития клетки-предшественники мигрируют из тимуса в лимфоидную ткань пищеварительного тракта. После рождения микроорганизмы, попавшие в желудочно-кишечный тракт новорожденного, контактируют со специфическими предшественниками Т-супрессоров и способствуют их дифференциации в Т-супрессорные клетки. Эти клетки и обеспечивают толерантность к тем микроорганизмам, которые внутриутробно индуцировали у плода формирование клеток-предшественников. Транзиторные микроорганизмы, к которым внутриутробно предшественники Т-супрессоров не сформировались, индуцируют ответ макроорганизма по типу первичного иммунного ответа с синтезом IgA, который предотвращает адгезию этих микроорганизмов и элиминирует их из организма.
Важное значение в индукции толерантности к нормальной микрофлоре принадлежит также адаптивному периоду в первые дни жизни новорожденного.
Таким образом, в зависимости от состава микрофлоры беременной женщины (нормоценоз или состояние дисбиоза) и функционального состояния фето-плацентарной системы происходят формирование микрофлоры новорожден-
Генитальные инфекции и факторы противоинфекционной защиты в генезе
воспалительных заболеваний женских половых органов 33
ного (колонизация представителями нормальной микрофлоры или УПМ) и становление его иммунной системы. Изучение этого вопроса только начинается, однако имеющиеся сведения уже позволяют считать важнейшей задачей нормализацию микрофлоры у беременных женщин для профилактики анте-и постнатальных инфекций.
В родах происходит механическое вымывание микроорганизмов из влагалища в момент излития околоплодных вод, и ребенок рождается в среде, содержащей небольшое количество бактерий с низкой вирулентностью.
В первые дни физиологического послеродового периода влагалище может содержать минимальное количество микроорганизмов. Лохии, обладая бактерицидным действием, вымывают микроорганизмы из влагалища. Однако вскоре происходит его заселение микроорганизмами с кожи промежности и из кишечника (стафилококки, колиформные бактерии, энтерококки), количество которых не превышает умеренных значений. При нормальном течении беременности и родов к 20-му дню послеродового периода заканчивается эпи-телизация эндометрия, а к 6-й неделе выделения из матки прекращаются. За это время происходит заселение влагалища лактобациллами и возвращение микрофлоры к исходному естественному состоянию.
1.3.1.4. Микрофлора влагалища в постменопаузе
С угасанием функции яичников уменьшается секреция всех половых гормонов, в том числе эстрогенов. На фоне дефицита эстрогенов эпителий влагалища становится тонким, так как прекращается митотическая активность клеток парабазального слоя и, следовательно, не происходит пролиферации вагинального эпителия, резко снижается синтез гликогена. Параллельно со снижением уровня гликогена из вагинального биотопа постепенно элиминируются лактобациллы. Распространено мнение о частой колонизации влагалища в этот период «фекальной» микрофлорой — энтерококками, эшерихиями и другими колиформными бактериями (Шендеров Б.А., 1998; Onderdonk A.B., Wis-semann K.W., 1993), что, в свою очередь, является фактором высокого риска возникновения воспалительных процессов генитального тракта в этом возрасте и развития восходящей мочевой инфекции (Alsina C.J., 1996). В постменопаузе ведущее место среди вагинальных инфекций занимает бактериальный вагиноз. Однако отсутствуют исследования, подтверждающие эту точку зрения на основе микроэкологических исследований, отвечающих современным требованиям, т.е. с количественной оценкой видового состава микроценоза.
При обследовании 57 женщин в возрасте от 47 до 71 года с длительностью постменопаузы от 1 до 19 лет проведена оценка микроэкологии влагалища на основе комплексного микробиологического исследования, включавшего куль-туральное исследование с определением количества и видового состава вагинальной микрофлоры и микроскопию вагинальных мазков, окрашенных по Граму, для выявления особенностей эпителия влагалища и характеристики микрофлоры по ее морфологическим особенностям (Онищенко Г.Г., Алеш-кин В.А., Афанасьев С.С. и др., 2002).
Анализ полученных результатов показал, что состояние микроэкологии влагалища определяется длительностью постменопаузы. У пациенток с длительностью постменопаузы от 1 года до 5 лет (1-я группа) выявлены 3 варианта вагинальной микроэкологии. У большинства женщин (11 из 18) диагностирован нормоценоз: отмечена умеренная общая микробная обсемененность вагинального отделяемого с абсолютным преобладанием лактобацилл (6—7 lg КОЕ/мл). При этом у 35% женщин лактобациллы находились в ассоциации с факульта-
34 Глава I
тивно анаэробными условно-патогенными бактериями (коагулазоотрицатель-ные стафилококки, энтерококки, коринебактерии), титр которых был низким (менее 3 lg КОЕ/мл). Вагинальный эпителий у этих женщин был представлен поверхностными и промежуточными клетками. У 6 пациенток этой группы был диагностирован бактериальный вагиноз и у одной — неспецифический вагинит, вызванный стрептококком группы В (5 lg КОЕ/мл) при отсутствии лактобацилл и умеренно выраженной лейкоцитарной реакции в вагинальном отделяемом.
Во 2-й группе женщин (20 пациенток) с длительностью постменопаузы от 7 до 10 лет почти с одинаковой частотой диагностированы нормоценоз (у 45%) и вагинальная атрофия (у 50%). При нормоценозе титр лактобацилл составлял 6—7 lg КОЕ/мл, у всех женщин лактобациллы были в ассоциации с УПМ, концентрация которых часто достигала умеренных значений (4—6 lg КОЕ/мл). Для вагинальной атрофии было характерным преобладание эпителиальных клеток парабазального и промежуточного слоев, скудная по количеству микрофлора (2—4 lg КОЕ/мл), представленная факультативно анаэробными условно-патогенными бактериями (эпидермальный стафилококк, колиформные бактерии, стрептококки групп В и D, коринебактерии).
В 3-ю группу вошли 19 женщин с длительностью постменопаузы 10—19 лет. Частота вагинальной атрофии в этой группе составила 73,7%. У остальных женщин состояние микроценоза влагалища было близким к нормоценозу, хотя имелись отличия от нормальных показателей репродуктивного возраста: титр лактобацилл был умеренным (5—6 lg КОЕ/мл) и в 100% случаев обнаруживались разнообразные виды условно-патогенных бактерий, титр которых также не превышал умеренных значений. При микроскопии вагинальных мазков всегда выявлялись парабазальные клетки при значительном снижении количества эпителиальных клеток поверхностных слоев.
Таким образом, проведенное исследование показало, что по мере старения женщины и прогрессирования эстрогенного дефицита происходят изменения микроэкологии вагинального биотопа. У большинства женщин с длительностью постменопаузы до 5 лет состояние микроэкологии влагалища соответствует показателям нормы репродуктивного периода, а его инфекционная патология представлена бактериальным вагинозом и неспецифическим вагинитом — заболеваниями, характерными для детородного возраста. При увеличении длительности постменопаузы до 10—19 лет у 50—74% женщин имелись проявления атрофии влагалища. При этом общее количество микроорганизмов в его отделяемом резко снижается (до 2—4 lg КОЕ/мл), а доминирующими становятся микроорганизмы условно-патогенной группы.
Дата добавления: 2014-12-16; просмотров: 1738;