Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА. и НШ. Клетка HI отличается от остальных длинными мощными дендритами, входящими в контакт с колбочками при помощи «триад»
 и НШ. Клетка HI отличается от остальных длинными мощными дендритами, входящими в контакт с колбочками при помощи «триад». Их аксон контактирует с палочкой. При этом образуются так называемые «точечные» синапсы [592, 597, 600]. Аксон также образует синапсы на дендритах биполярных клеток палочек.
и НШ. Клетка HI отличается от остальных длинными мощными дендритами, входящими в контакт с колбочками при помощи «триад». Их аксон контактирует с палочкой. При этом образуются так называемые «точечные» синапсы [592, 597, 600]. Аксон также образует синапсы на дендритах биполярных клеток палочек.
Приведенная выше схема синаптических связей является основой обработки информации, получаемой от многочисленных палочек. При этом информация собирается по площади сетчатки, превышающей площадь дендритного поля клетки.
Клеток типа НШ на 30% больше, чем типа HI, и они контактируют с колбочками.
Клетки типа НИ имеют тонкие дендриты и короткие (100—300 мкм) аксоны. Как дендриты, так и аксоны контактируют только с колбочками.
Недавние электронномикроскопические исследования сетчатки человека показывают, что существуют определенные закономерности в контакте между колбочками различных спектральных характеристик и различными типами горизонтальных клеток [45, 46, 230, 391] (рис. 3.6.26). Необходимо отметить, что клетки типа HI контактируют с колбочками всех спектральных типов. Наименее часто они контактируют с коротковолновыми («синими») колбочками. Клетки типа НИ, наоборот, чаще контактируют именно с «синими» колбочками, а клетки НШ с колбочками вообще не контактируют [597]. На основании этих данных предполагают, что клетки типа HI можно рассматривать как клетки «яркости», а клетки типов НИ и НШ как клетки воспринимающие цвета.
Горизонтальные клетки млекопитающих характеризуются также наличием многочисленных «щелевых контактов» между дендритами соседних клеток [594]. Благодаря этим контактам сигнал распространяется в плоскости синцития нейронов сетчатки. Помимо электрического сигнала через эти контакты могут проходить и низкомолекулярные вещества.
Строение тела горизонтальных клеток различных типов схоже. Тело клетки обычно уплощено и имеет диаметр 6—8 мкм. Ядро круглое и окружено аппаратом Гольджи. Цитоплазма содержит гладкую и шероховатую эндоплаз-матическую сеть, четкие митохондрии и многочисленные свободные рибосомы. Характерной особенностью горизонтальных клеток является наличие в цитоплазме включений, так называемых телец или «кристаллоида Колмера», описанного Колмером еще в 1918 г. [602]. Эти образования имеют длину 8—20 мкм и ширину 0,3—1,5 мкм и чаще обнаруживаются вблизи ядра, но видны и в цитоплазматических отростках [1119, 1212]. Состоят они из пакетов параллельно расположенных плотных трубочек в количестве от 5 до 30, отделенных промежутком шириной 2—6 мкм. Каждая трубочка
складывается из 2—3 концентрических мембран, на внутренней и внешней поверхностях которых лежат рибосомоподобные частицы, чувствительные к рибонуклеазе [1008]. Предполагают, что эти образования представляют собой своеобразную форму шероховатой эндо-плазматической сети. Функции горизонтальных клеток разнообразны. Более подробно о них будет изложено в 4-й главе. Здесь мы остановимся лишь на некоторых из них.
Во-первых, необходимо указать, что горизонтальные клетки интегрируют сигналы, поступающие от палочек и колбочек с выделением так называемых «каналов» передачи информации различного типа. При этом именно на уровне горизонтальных клеток уже четко определяется формирование структурно-функциональных нейронных единиц — «рецептивных полей» (см. главу 1 и 4), имеющих фундаментальное значение в обработке зрительной информации и передаче ее более высоко расположенным отделам центральной нервной системы. Именно благодаря «рецептивным полям» и формируются основные физиологические характеристики зрительного восприятия, такие как «контрастность», «цветовое зрение» и др.
Во-вторых, на основании выявления нейронных связей между горизонтальными клетками и фоторецепторами, а также физиологических исследований установлено, что горизонтальные клетки посылают зрительную информацию через синапсы обратной связи назад к фоторецепторам. Эти обратные связи способствуют функционированию «рецептивных полей».
В-третьих, благодаря наличию избирательных многоконтактных обратных связей горизонтальных клеток с палочками и колбочками различных спектральных характеристик, именно горизонтальные клетки объединяют и обрабатывают весь широкий спектр цветовой информации.
Биполярные клетки (рис. 3.6.27, 3.6.28). Биполярные клетки являются вторым нейроном зрительного пути. В каждой сетчатке содержится приблизительно 35 676 000 подобных клеток [137].
Тела этих клеток располагаются во внутреннем ядерном слое, а их отростки распространяются на наружный и внутренний плексиформ-ные слои.
Диаметр тела клетки в области желтого пятна равен 9 мкм, а в периферических отделах сетчатой оболочки — 5 мкм. В зависимости от типа синаптических отношений с другими клетками различают 9 основных типов биполярных клеток [138, 171, 600, 601, 693]. Восемь типов клеток относятся к биполярным клеткам колбочек и один тип к биполярным клеткам палочек. Это следующие типы:
1. Биполярные клетки палочек (щеткопо-
добные).
2. Инвагинированные карликовые.
Сетчатка
 | |||
 | |||
|
|
| BB |
| DBl FMB DB2 DB3 DB4 DB' |
| DB6 |
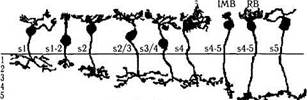
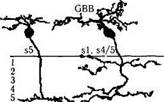
Рис. 3.6.27. Основные типы биполярных клеток сетчатки человека (по Kolb, I998):
DB— клетки диффузного типа; MB — карликовые клетки; ВВ — клетки «синих» колбочек; GBB — гигантские двухслойные; RB — биполярные клетки палочек. Приведенные слева цифры указывают уровень распространения дендритов клеток во внутреннем плексиформном слое

Рис. 3.6.28. Ультраструктурные особенности синап-тических контактов биполярных клеток на уровне внутреннего плексиформного слоя (по Kolb, 1998):
I — амакриновая клетка; 2 — биполярная клетка; 3—ганглиоз-
ная клетка. Кружками указаны места формирования синапсов
между различными типами клеток
3. Плоские карликовые.
4. Плоские диффузные.
5. Инвагинированые диффузные.
6. Биполярные клетки «синих» колбочек,
образующие ON-центр «рецептивные поля».
7. Биполярные клетки «синих» колбочек,
образующие OFF-центр «рецептивные поля».
8. Гигантские двухслойные.
9. Гигантские диффузные инвагинированные.
Биполярные клетки палочек (щеткопо-
добные) составляют 20% от общего числа биполярных клеток. Расположены они на расстоянии 1 мм от желтого пятна. Диаметр дендритного дерева клеток увеличивается по мере приближения клеток к периферии сетчатки
[600]. В наружном плексиформном слое основной дендрит клетки делится на 2—3 ветви, которые после прохождения между «ножками» колбочек образуют в виде щеточек отростки, проникающие в сферулу палочки.
В центральных участках сетчатки дендритное поле горизонтальных клеток маленькое (15 мкм) и дендриты контактируют с 15—20 палочками. По периферии сетчатки дендритное поле больше (до 30 мкм) и клетка входит в контакт с 40—50 палочками.
Аксоны биполярной клетки палочки во внутреннем плексиформном слое образуют синапсы с отростком амакриновой клетки, дендритами и телами клеток диффузных ганглиозных клеток (рис. 3.6.23, 3.6.28).
Плоские карликовые клетки самые маленькие. Дендриты клеток, имеющие вид пучка, проникают в «триаду» «ножек» колбочек. Апикальный дендрит экстрафовеолярных карликовых биполярных клеток делится на две части. При этом он образует синапсы с двумя различными колбочками. Аксоны переходят через внутренний плексиформный слой и образуют синапсы с отростками амакриновых клеток и дендритами «карликовых» ганглиозных клеток (рис. 3.6.28). В области центральной ямки одна карликовая биполярная клетка контактирует с одной колбочкой [600]. Эти биполярные клетки участвуют в образовании OFF-центр «рецептивных полей» колбочковой системы.
Плоские диффузные и инвагинированные «карликовые» биполярные клетки обладают многочисленными дендритами, заканчивающимися на «ножках» многих колбочек. Апикальный дендрит этих клеток разветвляется в наружном плексиформном слое, распространяясь в горизонтальной плоскости. Кроме того, эти биполярные клетки формируют обширную сеть в перифовеолярной области [600]. Инвагниро-ванные «карликовые» биполярные клетки участвуют в формировании ON-центр «рецептивных полей» колбочковой системы.
Биполярные клетки «синих» колбочек образуют синапсы более чем с одной «ножкой» колбочек [171, 425, 600]. Биполярные клетки «синих» колбочек чаще встречаются в 4 мм от желтого пятна, а их аксональные терминалы простираются до 30 мкм. Эти клетки имеют также два мощных дендрита, которые заканчиваются на той же самой колбочке или на другой колбочке или в нейропиле наружного плексиформного слоя.
Различают два типа гигантских биполярных клеток. Это деление определяется протяженностью дендритов клеток. В центральных участках сетчатки длина дендритов равна 50 мкм, а по периферии 100 мкм [600]. Биполярная клетка подобного типа объединяет 15—20 колбочек.
Гигантская диффузная биполярная клетка имеет толстый дендрит, который делится на
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 три длинные ветви, а также раздваивающийся аксон. Аксон заканчивается в 4-м слое внутреннего плексиформного слоя. Если бы не размер дендритного поля, гигантские диффузные биполярные клетки имели бы строение аналогичное строению плоской диффузной биполярной клетки.
три длинные ветви, а также раздваивающийся аксон. Аксон заканчивается в 4-м слое внутреннего плексиформного слоя. Если бы не размер дендритного поля, гигантские диффузные биполярные клетки имели бы строение аналогичное строению плоской диффузной биполярной клетки.
Все типы биполярных клеток близки по ультраструктурной организации. Ядро их круглое или овальное с одним или двумя ядрышками. Аппарат Гольджи, как и центриоли, располагается на участке выхода большого дендрита. Выполняют цитоплазму также рибосомы, шероховатый эндоплазматический ретикулум, митохондрии. В дендритах (толщина 0,1—0,2 мкм) также обнаруживаются митохондрии и микротрубочки, пузырьки и микрофиламенты (диаметр 20 нм). Аксональный бугорок расположен напротив выхода дендрита. В аксонах биполярных клеток выявляются и нейротрубочки (диаметр 12,5 нм). До внутреннего плексиформного слоя аксоны окружены отростками мюллеров-ских клеток.
После потери глиальной оболочки аксон образует утолщение (телодендрон), содержащее большое количество синаптических пузырьков, особенно вокруг синаптической ленты. Эфферентный или постсинаптический телодендрон обладает обычными синапсами, в то время как эфферентные отростки, образующие пресинап-тический контакт с амакриновыми и ганглиоз-ными клетками, обладают типичными ленточными синапсами.
Необходимо указать и на то, что основная часть внутреннего промежуточного слоя внутреннего ядерного слоя занята телами мюлле-ровских клеток, хотя они могут быть обнаружены и в любом другом слое сетчатки. Более подробно строение мюллеровской клетки изложено ниже.
Амакриновые клетки (рис. 3.6.29—3.6.31). Амакриновые клетки представляют собой нейроны, которые взаимодействуют на втором уровне вертикального пути передачи зрительной информации, а именно в направлении: фоторецептор — биполярная клетка — ганглиоз-ная клетка. Они формируют синапсы во внутреннем плексиформном слое. Эти клетки объединяют, а затем первично обрабатывают поступающую от биполярных клеток информацию и передают ее ганглиозным клеткам [39, 226, 228]. Тела амакриновых клеток находятся несколько кнутри от ядер клеток Мюллера. Каждая амакриновая клетка имеет единственный отросток, обладающий свойствами дендрита и аксона. Отростки распространяются в обширной области во внутреннем плексиформном слое.
Тело амакриновой клетки имеет форму колбы диаметром 12 мкм. Располагаются они во внутреннем ядерном слое за исключением области желтого пятна. Цитоплазма содержит мно-
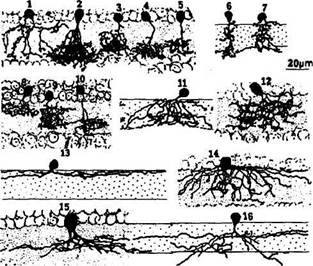
Рис. 3.6.29. Особенности распределения отростков
амакриновых клеток во внутреннем плексиформном
слое (объяснение в тексте)
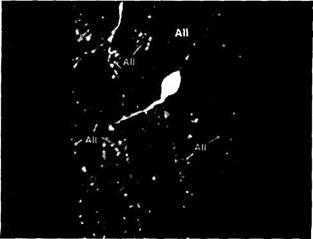

Рис. 3.6.30. Амакриновые клетки сетчатки, дающие положительную иммунногистохимическую реакцию, выявляющую серотонин (а) и допамин (б) (по Kolb, /995)
Сетчатка

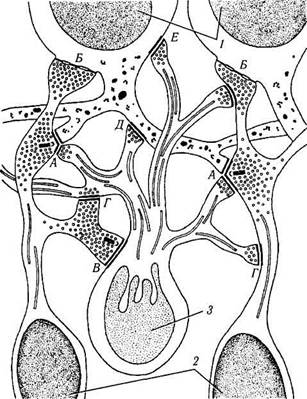
Рис. 3.6.31. Схематическое изображение синаптичес-
ких контактов между биполярными, амакриновыми и
ганглиозными клетками во внутреннем плексиформном
слое (по Hogan et al., 1971):
I — ганглиозные клетки; 2 — биполярная клетка; 3 — амакрино-вая клетка; А — аксодендритное окончание в диаде; Б — аксо-соматическое окончание на ганглиозной клетке; В — контакт между амакриновой и биполярной клетками; Г—аксоаксонный контакт между отростками амакриновой и биполярной клетками; Д — аксодендритный контакт между амакриновой и ганглиозной клетками; £—аксосоматический контакт между отростком амакриновой клетки и ганглиозной клеткой
гочисленные митохондрии, шероховатую эндо-плазматическую сеть (вещество Ниссля) и множество липидных включений. На внутренней поверхности клетки недалеко от ядра расположена ресничка.
В сетчатке человека амакриновые клетки отличаются разнообразным строением, и их описано 24 типа [154, 600]. При импрегнации сетчатки серебром по Гольджи выделяют два главных типа клеток: 1) диффузные и 2) стратифицированные.
Главный отросток клеток диффузного типа распространяется через все слои внутреннего плексиформного слоя. На его внутренней поверхности отросток разветвляется, формируя плотное горизонтально расположенное сплетение. В зависимости от протяженности отростков диффузные амакриновые клетки подразделяются на «узкопольные», охватывающие область шириной 10—50 мкм (составляет в среднем 25 мкм) и «широкопольные». Последние клетки во внутреннем плескиформном слое
распространяются на 30—50 мкм, а в слое ган-глиозных клеток до 600 мкм.
«Широкопольные» диффузные амакриновые клетки вступают в контакт с терминалами биполярных клеток палочек и ганглиозных клеток.
В зависимости от уровня расположения отростков во внутреннем плексиформном слое амакриновые клетки можно подразделить на следующие типы: нестратифицированные, муль-тистратифицированные и диффузные. Внутренний плексиформный слой еще Кахалом был условно подразделен на 6 слоев (страты). Это подразделение на слои используется морфологами для классификации амакриновых клеток до настоящего времени (рис. 3.6.29). Нестратифицированные амакриновые клетки лежат во внешней половине внутреннего плексиформного слоя и отдают отростки, длиной до 500 мкм.
Отростки мультистратифицированных клеток разделяются на ветви, простирающиеся на расстояние до 400—600 мкм. При этом они занимают два или более уровней во внутреннем плексиформном слое. Ядра стратифицированных диффузных клеток меньше, чем ядра других амакриновых клеток, а их отростки охватывают область, шириной не более 50 мкм.
Амакриновые клетки также можно классифицировать по обнаруживаемому в них типу нейромедиаторов. Нейромедиаторами этих клеток являются нейроактивные вещества (ацетил-холин, гамма-аминомасляная кислота (ГАМК), глицин, допамин, серотонин) и нейропептиды (холецистокинин, энкефалин, глюкагон, нейро-тензин, соматостатин, вещество Р, нейропеп-тид Y и вазоактивный кишечный пептид). В одной клетке могут присутствовать два или более перечисленных медиатора. Большинство амакриновых клеток содержат ГАМК, глицин, серотонин и допамин [154, 219, 237] (рис. 3.6.30).
Физиологическое значение амакриновых клеток интенсивно изучается в последние годы. Именно благодаря одновременному использованию морфологических, иммуногистохимических и электрофизиологических методов исследования выявлен ряд функций этих клеток. Получены эти данные в экспериментах на животных, в частности на кошках (см. главу 4).
У кошек различают несколько типов амакриновых клеток, функции которых достаточно хорошо изучены. Это амакриновая клетка А2, АН, А8, А13, А17, А19, А20, А22 и др. Об их роли будет рассказано в главе 4, посвященной зрительному пути.
Межплексиформные клетки. Межплекси-формные клетки описаны Gallego в 1971 г. [366]. Ядра межплексиформных клеток занимают самую внутреннюю часть внутреннего ядерного слоя. Поскольку тела клеток располагаются между амакриновыми клетками, некоторые авторы не выделяют этот тип клеток. Тем не менее отростки межплексиформных клеток, в
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 отличие от амакриновых, простираются на плексиформные слои [600]. Отростки к наружному плексиформному слою отходят непосредственно от тела межплексиформнои клетки или от их отростков. Сенсорный вход к межплек-сиформную клетку обнаруживается во внутреннем плексиформном слое, а большинство синапсов располагается в наружном плексиформном слое. Таким образом, информация передается между двумя указанными слоями. Межплексиформные клетки по своей природе относятся к центрифугальным нейронам. В сетчатке человека синапсы обнаруживаются, в основном, между межплексиформными клетками и горизонтальными клетками колбочек, получая при этом информацию от отростков ама-криновой клетки [661]. В зависимости от используемого нейромедиатора межплексиформные клетки позвоночных разделяют на три типа [526]. Это клетки, использующие ГАМК, тирозин гидроксилазу и глицин. У человека выявлен только 1 тип клеток (ГАМК) [154, 219].
отличие от амакриновых, простираются на плексиформные слои [600]. Отростки к наружному плексиформному слою отходят непосредственно от тела межплексиформнои клетки или от их отростков. Сенсорный вход к межплек-сиформную клетку обнаруживается во внутреннем плексиформном слое, а большинство синапсов располагается в наружном плексиформном слое. Таким образом, информация передается между двумя указанными слоями. Межплексиформные клетки по своей природе относятся к центрифугальным нейронам. В сетчатке человека синапсы обнаруживаются, в основном, между межплексиформными клетками и горизонтальными клетками колбочек, получая при этом информацию от отростков ама-криновой клетки [661]. В зависимости от используемого нейромедиатора межплексиформные клетки позвоночных разделяют на три типа [526]. Это клетки, использующие ГАМК, тирозин гидроксилазу и глицин. У человека выявлен только 1 тип клеток (ГАМК) [154, 219].
Внутренний сетчатый (плексиформный) слой(рис. 3.6.1, см. цв. вкл.; 3.6.31). Во внутреннем плексиформном слое контактируют второй (биполярная клетка) и третий (ганглиозная клетка) нейроны сетчатки. В пределах этого же слоя также взаимодействуют амакриновые и межплексиформные клетки. Кроме синаптичес-ких связей между биполярными, ганглиозными, амакриновыми и межплексиформными клетками этот слой содержит отростки мюллеровских клеток, а также обильную сосудистую сеть и ядра единичных ганглиозных и амакриновых клеток.
Внутренний плексиформный слой толще наружного плексиформного слоя. Отсутствует он только в области желтого пятна.
Во внутреннем плексиформном слое видны многочисленные синапсы, плотность которых достигает 2 млн в мм2 [263].
Существуют определенные ультраструктурные особенности этого слоя сетчатки. Именно в этом слое биполярные клетки вступают в си-наптический контакт с отростками амакриновых клеток и дендритами ганглиозных клеток, образуя так называемую «диаду». Наиболее часто один из элементов «диады» представляет собой дендрит ганглиозной клетки, а другой — отросток амакриновой клетки. Подобный тип синаптичекой организации соответствует наличию в этой области так называемых ганглиозных клеток «контрастности».
Реже в «диаде» обнаруживается два отростка амакриновой клетки или, что более редко, два дендрита ганглиозной клетки (рис. 3.6.31). Отростки амакриновых клеток связываются с аксонами биполярной клетки, телами и дендритами ганглиозных клеток посредством синапсов обычного строения. Межплексиформные клетки также образуют обычные синапсы, главным образом, с отростками амакриновых клеток.
В этом слое существует два уникальных типа синапсов, свойственных только амакриновым клеткам. Это «реципроктный» и «последовательный» синапсы. В «реципроктном» синапсе отросток амакриновой клетки в «диаде» образует синапс с терминалом биполярной клетки, обеспечивая, таким образом, обратную связь между амакриновой и биполярными клетками около синаптической ленты. «Последовательный синапс» состоит из двух последовательно расположенных синапсов между двумя отростками амакриновых клеток, а третий синапс образуется с дендритом ганглиозной клетки, аксоном биполярной клетки или другим отростком амакриновой клетки. Эта сеть обеспечивает взаимодействие соседних амакриновых клеток.
Синапсы амакриновых клеток располагаются слоями. Так, в области желтого пятна обнаруживается только два слоя синапсов, а по периферии число их слоев достигает пяти [154, 605].
Слой ганглиозных клеток(рис. 3.6.1, см. цв. вкл.). Слой ганглиозных клеток состоит в основном из тел ганглиозных клеток. В этом слое обнаруживаются также отростки мюллеровских клеток, нейроглия и сосуды сетчатой оболочки. Ганглиозные клетки получают обработанные зрительные сигналы от предшествующих двух нейронов, обрабатывают их и передают в центральную нервную систему [39, 154].
Ганглиозные клетки по периферии сетчатки образуют один слой клеток. С височной стороны диска зрительного нерва выявляется 2 слоя клеток, а по краям желтого пятна 6—8 слоев. В центре желтого пятна и диске зрительного нерва ганглиозные клетки отсутствуют.
Толщина слоя ганглиозных клеток колеблется от 10 до 20 мкм в назальной части сетчатки до 60—80 мкм в области желтого пятна [137].
В сетчатке взрослого определяется от 0,7 до 1,5 млн ганглиозных клеток. Соседние ганглиозные клетки плотно прилегают друг к другу за исключением периферии сетчатки. Здесь расстояние между ними достигает 400 мкм. Каждая клетка имеет один аксон. Собираясь на внутренней поверхности сетчатки, аксоны покидают глазное яблоко и формируют зрительный нерв.
В кольце, опоясывающем желтое пятно сетчатки, которое находится на расстоянии 0,4— 2,0 мм от пятна, плотность ганглиозных клеток колеблется от 32 000 до 38 000 клеток в мм2 [223]. По периферии плотность ганглиозных клеток в назальном квадранте в три разе превышает таковую в темпоральном квадранте. Плотность клеток в верхнем квадранте превышает плотность клеток в нижнем квадранте на 60%. Отношение количества колбочек к количеству ганглиозных клеток колеблется от 2,9:1 до 7,5:1.
В слое ганглиозных клеток обнаруживается до 3% амакриновых клеток в центральных
Сетчатка

|
| >,'!П |
областях сетчатки, по периферии почти 80% [223]. Ганглиозные клетки больших размеров (диаметр от 10 до 30 мкм), круглой или овальной формы. В области желтого пятна размер клеток несколько меньше [39, 154].

|
В цитоплазме развита шероховатая эндо-плазматическая сеть (вещество Ниссля) и аппарат Гольджи (рис. 3.6.32). Обнаруживаются также диффузно распределенные фрагменты гладкой эндоплазматической сети, митохондрии, капельки липидов и пигментные гранулы. С возрастом отмечается увеличение количества зерен липофусцина. С этим связывают усиление желтизны макулярной области.
Рис. 3.6.32. Ультраструктурные особенности ганглиоз-ной клетки (по Hogan, 1966):
отмечается хорошее развитие шероховатой эндоплазматической сети (стрелки). Цитоплазма насыщена овальными пигментированными частицами, придающими сетчатке желтоватый цвет
Ганглиозные клетки обладают многочисленными нейрофиламентами, что позволяет легко отличать ганглиозные клетки от мюллеровских.
Ганглиозные клетки относятся к мультипо-лярным нейронам. Их дендриты распределяются в горизонтальной плоскости сетчатки, а также проникают во внутренний плексиформный слой. Их аксоны направляются к слою нервных волокон, где они ориентируются параллельно внутренней поверхности сетчатки (рис. 3.6.33).
Ганглиозные клетки классифицируют в соответствии с размером тел клеток, степенью развития отростков и их протяженности. Классифицируют их также по типу синаптической связи с амакриновыми и биполярными клетками.
В последние годы описано приблизительно 18 различных морфологических типов ганглиоз-ных клеток. Пока не совсем ясно, являются они только морфологической разновидностью основного типа клеток или различны и их функции [39, 600].


Рис. 3.6.33. Особенности строения тела ганглиозных
клеток и их дендритного поля в различных участках
сетчатой оболочки (по Polak, 1940):
а — область центральной ямки; б — область экватора; в — периферия
В глазном яблоке человека идентифицировано два основных типа клеток, обозначенных как клетки М (зонтикоподобные) и клетки Р. В свою очередь клетки Р подразделяются на два подкласса: Р1, или карликовые нейроны, и Р2 (рис. 3.6.34).
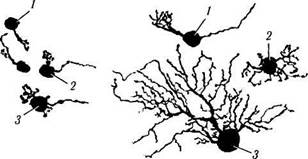
Рис. 3.6.34. Особенности строения ганглиозных клеток
сетчатки человека, определяющих функционирование
Р- и М-трактов зрительного анализатора:
/ — карликовая Р1; 2 — маленькая зонтикоподобная Р2; 3 — большая зонтикоподобная М
М-клетки проецируются на магноцеллюляр-ные (крупноклеточные) слои наружного коленчатого тела и определяют так называемый «не-оппонентный» ответ (см. главу 4). По своим
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 физиологическим характеристикам, М-клетки человека напоминают ганглиозные клетки обезьяны, специализированные на восприятии цвета.
физиологическим характеристикам, М-клетки человека напоминают ганглиозные клетки обезьяны, специализированные на восприятии цвета.
Р-клетки проецируются на парвоцеллюляр-ные (мелкоклеточные) слои наружного коленчатого тела. Клетки типа Р1 самые маленькие и обладают небольшим дендритным деревом.
Эти клетки определяют «оппонентный» ответ при стимуляции сетчатки средне- и коротковолновой частями спектра. Каждая Р-клетка получает информацию только от одной колбочки. Ганглиозные клетки типа Р1 подразделяют на так называемые широковетвистые (а-тип) и слабоветвистые (b-тип). Первые участвуют в формировании рецептивных полей с OFF-цент-рами, а вторые — с ON-центрами.
В области центральной ямки клетки типа Р1 составляют 90% общего количества ганглиозных клеток. В этой области сетчатки их удлиненные тела имеют размеры 8x12 мкм и достигают максимума (14x16 мкм) на расстоянии 8 мм от фовеа. Здесь они составляют 40—45% от общего количества ганглиозных клеток [227].
Единственный дендрит клетки формирует небольшое количество терминалов (5—7 нм в диаметре). Они переходят во внешнюю (а-тип) или внутреннюю (b-тип) треть внутреннего плексиформного слоя сетчатки [225, 595]. Синапсы а-типа образуются между аксонами плоских диффузных биполярных клеток, а синапсы b-типа — с инвагинирующими биполярными клетками.
В терминалах типа «диад» или «монад» присутствует до 55—81 лент. Количество отростков амакриновых клеток, которые образуют синапсы с дендритическим деревом этих ганглиозных клеток, приблизительно равно числу синап-тических лент биполярной клетки [595]. Распространяются дендриты ганглиозных клеток типа Р1 на 5—10 мкм в центральных областях сетчатки, а по периферии на 225 мкм [225, 592].
Отличить ганглиозные клетки Р1 от Р2 в области центральной ямки практически невозможно. Однако в 1,5 мм от нее клетки типа Р2 значительно больших размеров. Дендритное поле клеток, расположенных на расстоянии 6—8 мм от центральной ямки простирается на 30—50 мкм, а лежащих по периферии сетчатки на 400 мкм [225].
Клетки типа Р2 проявляют выраженный ответ при стимуляции светом S-колбочек [229]. Они составляют 1 % общего количества ганглиозных клеток в области фовеа и 10% по периферии сетчатой оболочки [227].
Размер М-ганглиозных клеток больше, чем клеток типа Р. Больше и их дендритные поля (25—30 мкм). Причем дендритное поле увеличивается по мере продвижения к периферии сетчатки. Так, в 8 мм от центральной ямки дендритное поле равняется 160 мкм, а на расстоянии 14 мм — 270 мкм [231].
М-клетки составляет 5% общего количества ганглиозных клеток в области центральной ямки и 20% по периферии сетчатки [227].
В литературе сейчас активно обсуждается вопрос о гибели ганглиозных нейронов сетчатки при глаукоматозном процессе, именуемом как глаукоматозная нейропатия. Само это состояние рассматривается как многолетний хронический процесс с постепенной медленной потерей отдельных ганглиозных клеток или их небольших групп при сохранении морфологии и функции других. Предполагалось, что в этом процессе преимущественно погибают магноцел-люлярные М-нейроны [851]. Однако Morgan et al. (2000) обнаружили, что в сетчатке обезьян с экспериментальной гипертензией в одинаковом соотношении погибают и магно- и парвоцеллю-лярные нейроны. При этом клетки сморщиваются, так что объем М-нейронов достоверно уменьшается на 20%, а Р-нейронов — на 16%.
Слой нервных волокон(рис. 3.6.1, см. цв. вкл.). Слой нервных волокон образуется аксонами ганглиозных клеток (так называемые «центростремительные», или «приводящие» волокна), а также глиальными элементами, большим количеством капиллярных сосудов и цент-рифугальных (эфферентных) волокон.
Аксоны ганглиозных клеток образуют дуги, очерченные отростками мюллеровских и других глиальных клеток. Отдельные афферентные волокна имеют диаметр от 0,6 мкм до 2,0 мкм. Они содержат микротрубочки, митохондрии и гладкую эндоплазматическую сеть. В них происходит двухсторонний аксоплазматический поток двух типов — медленный и быстрый. Медленный поток (0,5—5 лш/день) несет высокомолекулярные белки, используемые для роста аксонов и их физиологической регенерации. Быстрый поток (10—2000 лш/день) обеспечивает функционирование синапсов путем поставки питательных веществ [305, 722, 796].
Механизмы, обеспечивающие аксонный транспорт, изучаются до сих пор. В соответствии с одной из теорий движение в направлении аксона обеспечивается движением цитоплазмы [796]. Другие авторы считают, что в этом процессе основную роль играют микротрубочки. В подтверждение правильности последнего предположения приводится факт прекращения транспорта после обработки клеток колхицином, разрушающим микротрубочки [276].
Аксоны ганглиозных клеток сетчатки остаются немиелинизированными до момента достижения ими решетчатой пластинки.
Аффрентные волокна радиально продвигаются параллельно внутренней пограничной мембране и сходятся в области диска зрительного нерва. Исключением являются аксоны, исходящие из ганглиозных клеток, расположенных непосредственно с височной стороны диска зрительного нерва. Волокна папилло-макулярного пучка распространяются дугообразно. Верхние
Сетчатка
 и нижние волокна отделены горизонтальным «швом», простирающимся от желтого пятна до крайней периферии сетчатки.
и нижние волокна отделены горизонтальным «швом», простирающимся от желтого пятна до крайней периферии сетчатки.
Наиболее толстым является слой нервных волокон у края диска зрительного нерва с назальной стороны (20—30 мкм). Толщина его уменьшается по мере приближения к зубчатой линии. Значительно точнее варианты изменения толщины слоя нервных волокон можно выявить при помощи лазерной офтальмоскопии [830]. Эти данные важны при установлении диагноза ряда заболеваний глаза, в частности глаукомы.
Папилло-макулярный пучок является наиболее тонкой частью слоя нервных волокон, расположенного вокруг диска зрительного нерва. Поскольку наибольшее количество аксонов собирается с назальной стороны диска зрительного нерва, они образуют возвышенность (сосок), выстоящую в стекловидное тело.
Центрифугальные волокна, берущие свое начало в центральной нервной системе, заканчиваются во внутреннем плексиформном слое или самой внутренней части внутреннего ядерного слоя. Обычно они образуют синапс с амакри-новыми клетками или стенками капиллярных сосудов. В последнем случае эти волокна обеспечивают вазомоторные функции и регулируют интенсивность кровообращения.
Внутренняя пограничная мембрана.Внутренняя пограничная мембрана образует самый внутренний слой сетчатки и располагается на границе со стекловидным телом. Она является единственной истинной мембраной сетчатки. В образовании внутренней пограничной мембраны участвует как сетчатка, так и стекловидное тело. Состоит мембрана из четырех элементов: 1) коллагеновые волокна и 2) протео-гликаны (главным образом, гиалуроновая кислота) стекловидного тела; 3) базальная мембрана; 4) плазматическая мембрана мюллеровских клеток, возможно, и других глиальных клеток сетчатки.
Базальная мембрана положительно окрашивается при проведении ШИК-реакции.
Электронномикроскопически установлено, что коллагеновые волокна стекловидного тела, погруженные в протеогликаны, вплетаются в базальную мембрану глиальных клеток.
В задних отделах сетчатки внутренняя пограничная мембрана достигает толщины 0,5— 2,0 мкм. Она продолжается непрерывным слоем до желтого пятна, где значительно утолщается [473]. Отсутствует она по краю диска зрительного нерва, переходя в базальную мембрану астроцитов зрительного нерва [67]. По периферии сетчатки мембрана переходит в базальную пластинку эпителия ресничного тела. При старении внутренняя пограничная мембрана утолщается и прерывается в области зубчатой линии.
Внутренняя часть внутренней пограничной мембраны называется еще стекловидной мемб-
раной стекловидного тела. Именно она и придает поверхности сетчатки характерный блеск, наблюдаемый при офтальмосокопии. Обычно стекловидное тело плотно прилежит к сетчатке у диска зрительного нерва, в области центральной ямки и у зубчатой линии.
3.6.3. Зрительные пигментыи фоторецепция
Описывая строение сетчатой оболочки, необходимо хотя бы кратко остановиться на процессах, происходящих в фотороцепторах и определяющих понятие фоторецепции.
Процесс восприятия света связан непосредственно с физико-химическими процессами, происходящими в стопках мембран наружных члеников палочек и колбочек, и представляет собой целую систему связанных между собой химических преобразований, направленных на трансформацию световой энергии в нервный импульс. В систему этих преобразований входят также механизмы, направленные на восстановление веществ, обеспечивающих световос-приятие, регенерацию наружных члеников фо-торецепторых клеток и др. Центральное место в восприятии световой энергии занимают специализированные вещества — зрительные пигменты, которые располагаются именно в мембранах наружных члеников фоторецепторных клеток.
Сейчас мы кратко остановимся на сути происходящих при световосприятии процессах. Первоначально мы опишем особенности химической организации мембран наружных члеников фотороцепторов и зрительных пигментов.
Как было указано выше, мембраны наружных сегментов палочек и колбочек содержат зрительные пигменты, которые абсорбируют световую энергию и инициализируют зрительное возбуждение. Эти белковые молекулы внедрены в двухслойные липидные мембраны пластин наружных члеников фоторецепторов (рис. 3.6.35, см. цв. вкл.). В наружных сегментах палочек липиды и белки составляют примерно 50% веса. Большинство липидов относятся к фосфолипидам. В состав фосфолипи-дов, помимо глицерина, входят также две цепи жирной кислоты (в положении 1 и 2) и фосфорнокислая группа (в положении 3). В липидном слое мембраны цепи жирных кислот ориентированы таким образом, что внутри мембраны образуется гидрофобная область, а снаружи располагаются глицерол/фосфатные группы, обеспечивающие гидрофильность этой поверхности мембраны. Характерной особенностью липидов сетчатки является высокое их насыщение ненасыщенными жирными кислотами.
Наружные сегменты палочек содержат также большое количество различных белков, главным из которых является опсин. Родопсин представляет собой соединение 11-цис-ретина-
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 ля (альдегид витамина А) с опсином посредством основания Шиффа. Родопсин относится к так называемым трансмембранным белкам, N-конец которых обращен в междисковое пространство, а С-конец обращен в цитоплазму (рис. 3.6.35) [447, 448]. Благодаря этому молекулы хромофора ориентированы параллельными рядами вдоль мембран наружных члеников фоторецепторов, т. е. располагаются перпендикулярно падающим на него фотонам, обеспечивая максимальный сбор световой информации. Установлено, что диск наружного сегмента палочки содержит от 300 до 900 молекул родопсина [447, 448].
ля (альдегид витамина А) с опсином посредством основания Шиффа. Родопсин относится к так называемым трансмембранным белкам, N-конец которых обращен в междисковое пространство, а С-конец обращен в цитоплазму (рис. 3.6.35) [447, 448]. Благодаря этому молекулы хромофора ориентированы параллельными рядами вдоль мембран наружных члеников фоторецепторов, т. е. располагаются перпендикулярно падающим на него фотонам, обеспечивая максимальный сбор световой информации. Установлено, что диск наружного сегмента палочки содержит от 300 до 900 молекул родопсина [447, 448].
В сетчатке человека выявлено четыре типа зрительных пигментов. Один тип обнаружен в палочках (родопсин) и три в колбочках (иодо-псин). В зависимости от спектральных особенностей поглощения световой энергии колбочко-вые пигменты разделяются на чувствительные к красной (570 нм), зеленой (540 нм) и синей частям спектра (440 нм). 11-цис-ретиналь является хромофором для всех четырех классов зрительных пигментов человека.
Основным механизмом преобразования световой энергии является изменение характера взаимодействия хромофора (11-цис-ретиналь) с белком (опсин). Механизм этого процесса сводится к тому, что при действии световой энергии происходит изомеризация 11-цис-ретиналя с превращением его в полностью транс-рети-наль (рис. 3.6.36). Изменение строения молекулы ретиналя разрушает ее связь с опсином, что приводит к нарушению третичной структуры белка. Этот процесс происходит через ряд звеньев с образованием промежуточных продуктов. Эти промежуточные вещества существуют
| 11 -цис-ретинил эфир |

|
| 11-цис-ретинол |
| ПЭС |
 Полностью-транс-ре-тинил эфир
Полностью-транс-ре-тинил эфир
| 11-цис ретинол__ НАдф НАДФН + Н+- |

|
| X та О ч |
| Полностью-транс-ретиналь 1 Опсин |


 Полностью-транс-ретинол
Полностью-транс-ретинол
11-цис ретиналь
Родопсин
| JI |

 НСП
НСП
Рис. 3.6.36. Химические превращения родопсина в процессе зрительного цикла:
ПЭС — пигментный эпителий сетчатки; НСП — наружный сегмент палочки
исключительно короткое время и их можно анализировать только при низких температурах (рис. 3.6.37). Наиболее важным звеном в этом процессе является переход метародопсина I в метародопсин II. Именно на этом этапе и происходят конформационные изменения белковой части родопсина, что приводит к появлению у последнего ферментативной активности. Эти изменения инициируют дальнейший каскад процессов преобразования, о которых речь пойдет несколько ниже [78, 371, 448].
Родопсин (498 нм)
Свет ------ *- I Пикосекунды
Прелюмиродопсин (батородопсин) (543 нм)
I Наносекунды
Люмиродопсин (497 нм)
I Микросекунды
Метародопсин I (478 нм)
I Миллисекунды
Метародопсин II (380 нм)
I Секунды
Метародопсин III (465 нм)
I Минуты
Опсин (280 нм)
+ Транс-ретиналь (380 нм)
Рис. 3.6.37. Схема превращений родопсина под действием световой энергии (в скобках указаны спектральные изменения продуктов реакции)
После разрушения связи хромофора с опсином наступает обратный процесс, т. е. регенерация родопсина. Происходит это следующим образом (рис. 3.6.36). При обесцвечивании зрительного пигмента полностью-транс-ре-тиналь высвобождается из зрительного пигмента и преобразуется в полностью-транс-ретинол. Полностью-транс-ретинол из наружных сегментов фоторецепторов поступает в пигментный эпителий сетчатки, где он эстерифицируется, превращаясь в эфир полностью-транс-ретинил эфир. Последний превращается в 11-цис-ретинол благодаря деятельности фермента — рети-ноид изомеразы. Образовавшийся в результате реакции 11-цис-ретонол возвращается в фоторецепторы, где, окисляясь, превращается в 11-цис-ретиналь. 11-цис-ретиналь соединяется с опсином, образуя родопсин. Вновь образованный родопсин может опять абсорбировать
Сетчатка

|
|
| Темновые условия Наружный сегмент Внутренний сегмент |
| 3Na* |
| -No* |
| Свет |
фотон и инициализировать зрительный цикл. Таким образом, та же самая молекула опсина может многократно использоваться в зрительном возбуждении.
Из приведенной цепи реакций видно, что составленные части родопсина повторно используются в зрительном цикле. Тем не менее процесс регенерации хроматофора предполагает обязательное постоянное пополнение клеток пигментного эпителия витамином А, из которого образуется эфир 11-цис-ретинила.
В организм человека витамин А поступает с пищей и хранится в печени. Поступая в кровь, он связывается с ретинол-связывающим белком и затем с преальбумином. Этот белковый комплекс, благодаря наличию фенестр в эндотели-альной выстилке капиллярных сосудов хориои-деи, легко проникает через мембрану Бруха и достигает клеток пигментного эпителия сетчатки. Затем витамин А отделяется от белковой части комплекса и поступает в цитоплазму пигментных клеток для дальнейших преобразований в 11-цис-ретиналь.
Для восстановления родопсина необходимо пополнение и его белковой части, т. е. опсина. Пополнение фоторецепторов опсином происходит благодаря постоянно протекающему процессу регенерации наружных члеников палочек и колбочек. Вновь образованные мембранные пакеты, содержащие в своем составе и опсин, постепенно передвигаются к апикальной поверхности фоторецептора, где опсин связывается с 11-цис-ретиналем, образуя «новый» родопсин.
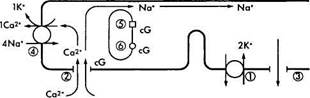
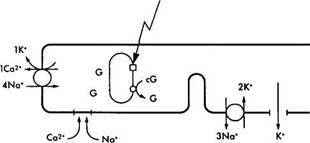
Кратко описав характер химических преобразований родопсина в процессе зрительного цикла, необходимо ответить на вопрос — каким образом описанные физико-химические процессы приводят к инициализации нервного импульса? Чтобы понять этот процесс необходимо обратиться к рис. 3.6.38. На рисунке видно, что в темновых условиях фоторецепторы деполяризованы. Это связано с тем, что натриевые каналы плазматических мембран сегментов палочек и колбочек в темноте открыты и из внеклеточного пространства в цитоплазму фоторецептора поступает большое количество ионов натрия. При этом диффузия натрия из наружного сегмента фоторецепторов во внутренний сегмент в темновых условиях обеспечивает формирование «темнового тока» [1206].
Натриевые каналы остаются открытыми благодаря высокой концентрации циклического гуанозин монофосфата (cGMP). Равновесие между ионами натрия и калия поддерживается благодаря деятельности АТФ-зависимого натрий/калиевого насоса.
Воздействие на зрительный пигмент световой энергии приводит к закрытию ионных каналов и снижению проводимости Na+ через мембрану наружного сегмента (рис. 3.6.38). При этом изменяется трансмембранный потенциал фоторецептора и возникает гиперполяризация.
Рис. 3.6.38. Схематическое изображение механизма
формирования нервного импульса в фоторецепторной
клетке:
В темноте ионы натрия (Na+), как и ионы кальция (Са+), перемещаются из наружного сегмента фоторецептора во внутренний благодаря деятельности Na+/K насоса (/), а поступают в наружный сегмент через катионные каналы (2). При этом формируется «темновой ток» ионов натрия. Катионные каналы открыты тогда, когда сСМР (cG) находится в связанном состоянии. Поток ионов натрия в направлении внутреннего сегмента происходит по мере выхода из клетки ионов калия (3). Вследствие поглощения фотона родопсином (5) активизируется фосфодиэстераза (6), что приводит к повышению концентрации cGNP и закрытию катионных каналов. Следствием этого является уменьшение проницаемости мембраны для ионов натрия и усиление ее поляризации. Посредником в этом процессе являются ионы кальция, поскольку они эффективно блокируют натриевые каналы и вызывают наблюдаемую гиперполяризацию. Выведение ионов кальция обеспечивается деятельностью ионообменника (4) и при закрытых каналах
Таким образом, фоторецепторы отвечают на освещение не потенциалами действия, а гиперполяризацией, величина которой пропорциональна интенсивности освещения.
В деполяризованном (темновом) состоянии фоторецепторы высвобождают нейромедиаторы в синаптическую щель, которые взаимодействуют с постсинаптическими терминалами биполярных и горизонтальных клеток.
Увеличение степени освещенности вызывает градуированную гиперполяризацию, которая вызывает уменьшение выделения нейромедиа-тора.
Необходимо отметить, что фоторецепторы, как и горизонтальные и биполярные клетки, не генерируют потенциалы действия, и таким образом отвечают на световую энергию уменьшением выделения медиатора [1206, 1055]. Только нейроны третьего порядка (ганглиозные клетки) генерируют потенциалы действия.
Как указано выше, индуцирует гиперполяризацию фоторецептора перекрытие ионных каналов. Изучению механизмов этого процесса
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

 посвящено большое количество исследований. Установлено, что основную роль в закрытии ионных каналов играет циклический гуанозин монофосфат (cGMP). Именно cGMP индуцирует целый каскад реакций ферментативного превращения различных белков с участием ионов кальция. Одну из центральных ролей в этом процессе играют белок-передатчик трансдуцин и фермент фосфодиэстераза. Именно фосфоди-эстераза снижает концентрацию cGMP, что и приводит к закрытию ионных каналов.
посвящено большое количество исследований. Установлено, что основную роль в закрытии ионных каналов играет циклический гуанозин монофосфат (cGMP). Именно cGMP индуцирует целый каскад реакций ферментативного превращения различных белков с участием ионов кальция. Одну из центральных ролей в этом процессе играют белок-передатчик трансдуцин и фермент фосфодиэстераза. Именно фосфоди-эстераза снижает концентрацию cGMP, что и приводит к закрытию ионных каналов.
Необходимо указать и на то, что фоторецептор не просто регистрирует световую энергию. Он также адаптируется к степени освещенности. Например, колбочки могут адаптироваться таким образом, что наша зрительная система регистрирует свет от слабых интенсивно-стей освещения до ярко освещенных солнцем объектов.
3.6.4. Нейромедиаторы
( нейротрансмиттеры ) сетчатой оболочки
Описывая микроскопическое строение сетчатой оболочки, мы неоднократно упоминали о наличии определенных структурных отличий синаптической организации нейронов сетчатки. Нейроны сетчатки отличаются и используемым типом нейромедиатора при передаче информации от одного нейрона другому.
В последнее время было обращено особое внимание на изучение нейромедиаторов сетчатки, что позволило более точно дифференцировать различные типы нейронов и выявить их функцию. Развитию направления изучения нейромедиаторов способствовали успехи смежных дисциплин, таких как ауторадиография, иммунология и молекулярная биология.
Клетки, окрашенные конъюгированными к различным типам нейромедиаторов антителами, меченными пероксидазой хрена, окрашивают самые нежные нервные волокна. На основании этого возможна довольно точная дифференциация клеток, особенно при одновременной их импрегнации по Гольджи. Большинство исследований нейромедиаторов нейронов сетчатки проведено на животных, но многое и на сетчатке человека [39, 219, 453]. Необходимо отметить, что полученные данные при исследовании животных во многом совпадают с данными исследования сетчатки человека.
Перед тем как более подробно остановиться на каждом из выявленных в сетчатке нейро-медиаторе, необходимо указать, что все они обнаруживаются и в центральной нервной системе, что еще раз доказывает существование единства механизмов их развития и функционирования.
Глютаминовая кислота.Глютаминовая кислота относится к наиболее распространенным нейромедиаторам нейронов «вертикальных»
нейронных трактов сетчатки (рис. 3.6.39, а). Все фоторецепторы используют глютаминовую кислоту для передачи сигналов к нейрону следующего порядка [237, 453, 702].
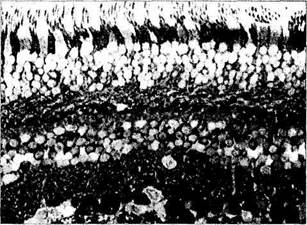
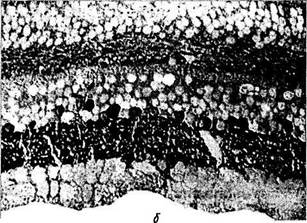
Рис. 3.6.39. Распределение глютаминовой кислоты (а) и гамма-аминомасляной кислоты (б) в нейронах сетчатой оболочки человека:
интенсивное черное окрашивание цитоплазмы клеток различных слоев сетчатки свидетельствует о положительной гистохимической реакции на выявляемый медиатор. Наиболее интенсивное окрашивание выявляется в цитоплазме ганглиозных клеток, менее интенсивное в нейронах внутреннего и наружного ядерных слоев (стрелки)
Предполагают, что глютаминовая кислота является нейромедиатором всех биполярных и большинства ганглиозных клеток сетчатки позвоночных [219, 691].
Поглощение, высвобождение и физиологическое действие глютамата и его агонистов на нейроны второго порядка подтвердило, что глютамат является нейромедиатором возбуждающего действия в первом синапсе сетчатки. Действие этого нейромедиатора на нейроны второго порядка происходит посредством двух различных типов сенсорных каналов. Один тип постсинаптического рецептора относится к ме-таботропному, а второй является ионотропным [771, 829]. Метаботропные рецепторы активи-
Сетчатка
 зируются посредством G-белка. Ионотропные рецепторы представляют собой интегральные мембранные белки, фиксирующие глютамино-вую кислоту. Этот процесс приводит к открытию катионных каналов. В настоящее время выявлен целый ряд ионотропных рецепторов [453, 847, 848].
зируются посредством G-белка. Ионотропные рецепторы представляют собой интегральные мембранные белки, фиксирующие глютамино-вую кислоту. Этот процесс приводит к открытию катионных каналов. В настоящее время выявлен целый ряд ионотропных рецепторов [453, 847, 848].
Дендриты биполярных клеток, расположенные в наружном плексиформном слое, имеют рецепторные каналы, которые относятся или к метаботропным или ионотропным. В то же время их аксоны, расположенные во внутреннем плексиформном слое, имеют каналы и рецепторы для гамма-аминомасляной кислоты (типов А, В и С), допамина и глицина. Это связано с тем, что все виды амакриновых клеток являются на этом уровне внутреннего плексиформного слоя пресинаптическими [154].
Гамма-аминомасляная кислота (ГАМК).Классический тормозной нейромедиатор, гамма-аминомасляная кислота (ГАМК), встречается во многих типах амакриновых и горизонтальных клеток у большинства позвоночных [691] Имеются некоторые противоречия при решении вопроса — содержится ли ГАМК в горизонтальных клетках обезьян и человека.
При окрашивании сетчатки человека видно, что четко окрашивается внутренний плекси-формный слой и приблизительно половина тел амакриновых клеток, лежащих во внутреннем ядерном слое. Горизонтальные клетки не окрашиваются (рис. 3.6.39, б).
Благодаря использованию двойных методов окрашивания стало известно, что амакриновые клетки типа А2, А10, А13, А17, А19 и меж-плексиформная клетка накапливают ГАМК и, вероятно, используют ее как первичный нейромедиатор. Некоторые амакриновые клетки одновременно с ГАМК используют и другие нейромедиаторы, такие как серотонин, ацетил-холин (звездчатые амакриновые клетки), допа-мин [1128], нейропептиды (вещество Р).
ГАМК-эргические амакриновые и межплек-сиформные клетки действуют на отростки биполярных, амакриновых и ганглиозных клеток или тела клеток в нейропиле сетчатки посредством всех трех типов ГАМК рецепторов.
Глицин.Глицин является аминокислотой. В центральной нервной системе и сетчатке глицин выполянет медиаторные функции. Определяется он в амакриновых клетках, не дающих реакцию на ГАМК [341, 691]. Предполагают, что к глицинэргическим относятся также несколько типов биполярных клеток. Глицин осуществляет некоторые формы постсинаптичес-кого торможения.
В сетчатке человека выявляется два морфологических типа глицинэргических амакриновых клеток. Менее интенсивно окрашиваются клетки типа АН. Более интенсивно окрашиваются клетки А4 и А8 [841, 842]. Глициновые рецепторы также найдены на всех нейронах,
являющихся постсинаптическими по отношению к амакриновым клеткам — на аксонах биполярных клеток, на дендритах ганглиозных клеток. Обнаружены глициновые рецепторы и в мюллеровских клетках [265].
Допамин.Нейромодулятор допамин обнаруживается в нескольких типах амакриновых клеток сетчатки млекопитающих. Наиболее интенсивно окрашивается при проведении иммуно-гистохимической реакции амакриновая клетка типа А18 [600].
Допаминовая клетка первого типа (А18)образует синапс на амакриновой клетке палочки АН и, возможно, также на клетках А8 и А17 [179, 591, 843, 1150].
Второй тип допаминовой амакриновой клетки был описан у обезьян и человека [694]. Эта клетка отдает дендриты, распределяющиеся в 3-м слое (страте) внутреннего плексиформного слоя.
Допаминовые клетки первого типа обеспечивают функционирование восходящих путей, направляющихся к наружному плексиформному слою. В этом слое они образуют синапсы с ГАМК-эргическими межплексиформными клетками.
Допаминовые рецепторы (D1 и D2) были идентифицированы на нейронах внутреннего и наружного ядерных слоев сетчатки многих позвоночных. Предполагают, что рецептор D1 характерен для горизонтальных клеток наружного плексиформного слоя и некоторых амакриновых клеток внутреннего плексиформного слоя. Рецепторы D1 также выявлены на телах ганглиозных клеток.
Рецепторы D2 найдены в наружном ядерном слое, наружной пограничной мембране и даже в пигментном эпителии сетчатой оболочки. Присутствуют они и во внутреннем плексиформном слое.
Ацетилхолин.Классический эксцитатный нейромедиатор периферической нервной системы — ацетилхолин. Он найден в амакриновых клетках сетчатки позвоночных. У кролика такие клетки были названы звездоподобными клетками [299, 700]. Различают два типа подобных клеток. Клетки одного типа располагаются в субпластинке авнутреннего плексиформного слоя. Тело другого типа клеток смещено к слою ганглиозных клеток, а их дендриты распределяются в субпластинке Ь.
Эти ацетилхолинсодержащие амакриновые клетки близки по строению почти у всех позвоночных. Описаны они и в сетчатке человека [514, 600].
Как мускариновые, так и холиномиметичес-кие рецепторы выявлены в сетчатке млекопитающих, особенно в ганглиозной клетке (Y-клет-ки). Влияние ацетилхолина и его антагонистов на ганглиозную клетку пока неясно.
Серотонин.Имеется два типа серотонин-содержащих амакриновых клеток в сетчатке
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 кролика [1128]. Один из них относится к клетке типа А17 системы палочек. В сетчатке кошки выявлены совершенно иные серотонинэрги-ческие типы амакриновой клетки. Первый тип клеток кошки подобен широкопольной клетке А20, а второй—А18 или дофаминовой клетке [1149].
кролика [1128]. Один из них относится к клетке типа А17 системы палочек. В сетчатке кошки выявлены совершенно иные серотонинэрги-ческие типы амакриновой клетки. Первый тип клеток кошки подобен широкопольной клетке А20, а второй—А18 или дофаминовой клетке [1149].
Аденозин. Пуриновый нуклеотид аденозин в сетчатке млекопитающих может быть нейро-медиатором или нейромодулятором. Ауторадио-графия и иммуногистохимия показали наличие аденозина в амакриновых и ганглиозных клетках [129]. В сетчатке человека также выявляются аденозинсодержащие клетки, которые можно отнести к биполярным и горизонтальным клеткам.
Влияние аденозина на сетчатку и на функции ганглиозных клеток верхних бугорков четверохолмия было зарегистрировано при использовании методов электрофизиологии, что подтверждает его нейромедиаторную роль [129].
Пептиды.В настоящее время на роль пептидных нейромедиаторов в ткани мозга претендует около 50 белков. Из них приблизительно четвертая часть выявлется в сетчатой оболочке. Это вазоактивный кишечный полипептид (VIP), вещество Р, энкефалины, соматостатин, нейроактивный пептид Y, глюкагон, холецисто-кинин и нейротензин. Перечисленные медиаторы выявлены в амакриновых клетках сетчатки разнообразных животных. Более подробно мы остановимся на веществе Р.
Вещество Р относится к нейропептидам. Состоит оно из 11 аминокислот и принадлежит к семейству тахикининов, включающему нейро-кинин А, нейропептид К и нейрокинин В. Вещество Р является нейромедиатором или нейромодулятором сетчатки млекопитающих [598].
| Рис. 3.6.40. Субстанция Р в амакриновых клетках сетчатки человека (по Kolb et al., 1995) |
Только среди амакриновых клеток сетчатки человека обнаруживаются Р-эргические клетки (рис. 3.6.40). Эти клетки отличаются широким дендритным полем, достигающим 3-го слоя (S3) внутреннего плексиформного слоя. Здесь от-
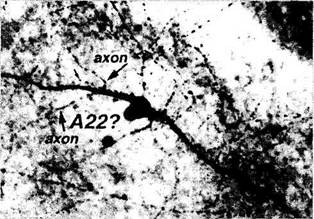
ростки формируют густое сплетение. Либо от тел клеток, либо от их дендритов отходят «аксон-подобные» отростки, которые, в свою очередь, разделяются на два длинных нежных отростка, расходящихся в противоположных направлениях на сотни микрон и заканчивающихся в слоях S5 и S3. Длинные дендриты этих клеток заканчиваются также на стенках кровеносных сосудов.
Оксид азота.Окись азота образуется во многих нейронах периферической и центральной нервной системы и выполняет нейромедиаторную роль. Косвенно способность клеткой синтезировать оксид азота можно выявить путем проведения гистохимической реакции, выявляющей активность НАДФ-диафоразы. При применении этого метода выявлено три типа амакриновых клеток и один тип ганглиозной клетки, дающих четкую реакцию на НАДФ-диафоразу. Эти клетки обладают большим телом и лежат в слое амакриновых клеток или смещены к слою ганглиозных клеток. Их дендриты достигают 3-го слоя внутреннего плексиформного слоя.
Дата добавления: 2014-12-20; просмотров: 1005;