Основи порівняльноїембріології
Ебріологія (від гр. ешьгіоп - зародок, 1о§оз —■ вчення) - наука про закономірності розвитку живих істот з моменту запліднення до утво-рення сформованого організму.
Ембріологію поділяють на загальну і спеціальну. Загальна ембріо-логія вивчає розвиток і будову статевих клітин (прогенез), запліднення, ранні етапи розвитку зародка, формування осьових органів і плодових оболонок (ембріогенез). Спеціальна ембріологія вивчає розвиток окре-мих органів до дефінітивного стану (органогенез). Первісні відомості про розвиток тварин набувалися людиною шляхом спостережень, ко-трі передавалися від покоління до покоління.
В давній Греції природознавство досягло високого рівня. Вели-кий вчений Гіпократ (народ. в 420 р. до н.е.) вперше описав розвиток зародка в курячому яйці. За його уявленням курча виникає із жовт-ка і живиться білком. Арістотель (народ. в 385 р. до н. е.) створив оригінальну класифікацію яєць різних тварин, обгрунтував функції плаценти та пуповини, описав процеси розвитку внутрішніх органів в ембріогенезі.
Прогресивний розвиток ембріології почався після створення світ-лооптичного мікроскопу. Так, на початку XVII ст. італійський вчений М. Мальпігі описав початкові стадії розвитку нервової пластинки, сер-ця, хребта та судин курчати.
Велике значення мало відкриття Р. Левенгуком в 1690 р. сперматозо-їдів. Він назвав їх «анімалькулі», тобто маленькі тварини. В цей же пері-од англійський вчений В. Гарвей описав кровообіг, а також встановив, що всі тварини і навіть нижчі (не досконалі) виникають шляхом розвитку із яйця. Його положення «все живе з яйця» (отпіа уіуо ех оуо) спростува-ло існуюче уявлення про мимовільне зародження живих істот.
Розділ 2 Ембргологія
До початку XVIII ст. в свідомості людей була ще дуже міцна ідео-логія феодального суспільства і загальним визнанням користувалося ідеалістичне вчення про преформацію (від лат. ргаегогтаге - заздале-гідь формувати). Згідно цьому вченню ніщо не виникає заново в світі живих тварин. Всі організми і всі їх органи не створюються в процесі розвитку а були закладені у вигляді нескінченно малих, але повністю сформованих зачатків ще при «створенні всесвіту». Тобто при виник-ненні організму не з'являється нового, а тільки ріст цих зачатків.
Преформісти поділялися на анімалькулістів, які вважали, що сфор-мований організм закладений у сперматозоїді, та овістів які здатністю нести сформований організм наділяли яйцеклітину.
В сформуванні ембріології в самостійну науку велике значення малипрацівидатнихвченихРосії. ЗокремаК. Ф. Вольф (1733-1794 рр.) в праці «Теорія генерації» показав, що процес ембріонального розвитку є не тільки збільшенням маси тіла, його ростом, але й новоутворенням. Він обгрунтував теорію епігенезу (від гр. ері - над, §епезіз — розвиток).
Подальший розвиток ембріології пов'язаний з іменами академіків X. I. Пандера та К. М. Бера. Першому з них належить честь відкриття і описання бластодерми і зародкових листків у ембріона курчати, отри-мання нових даних про розвиток травної та кровоносної систем.
К. М. Бер ( 1792 - 1876 рр.) є одним з творців порівняльної ембрі-ології. Він вперше виявив та описав яйцеклітини ссавців. Вчений до-сліджував яєчники собак, корів, свиней, кролиць, їжачих, дельфінів, жінок. В граафових пухирцях він виявив яйцеклітини. 3 нагоди його 50-річчя від Петербурзької Академії Наук К. М. Беру була вручена ювілейна медаль з надписом «оіїліз аБ оуо потіпет озіепсііі» (почав з яйця він показав людині людину).
Друга половина XIX століття позначилася знаменитими дослі-дженнями О.О.Ковалевського та 1,1. Мечнікова. На основі величезно-го фактичного матеріалу їм вдалося довести, що ранні етапи розвитку безхребетних та вищих хребетних тварин відбуваються однотипно. Еволюційна теорія підтвердила єдність усього тваринного світу.
О.О. Ковалевський і І.І. Мечніков заклали основи нової науки — порівняльної ембріології, яка дала необхідні факти еволюційній теорії.
Одним із важливих напрямів еволюційної ембріології є експери-ментальна ембріологія, основи якої були закладені О.М. Северцевим, П.П. Івановим, Д.П. Філатовим, Г.А. Шмідтом та іншими вченими.
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
Розвиток сучасної ембріології тісно пов'язаний із застосуванням нових методів дослідження, серед яких важливе місце належить гісто-хімії, електронній мікроскопії, гістоавторадіографії, культурі тканин, мікрургії, морфометрії та ін.
Знання ембріології та методи мають безпосереднє значення для культивування яйцеклітин і зародків та імплантації їх в матку; при створенні відповідних умов, необхідних для нормальної життєдіяль-ності клітин, тканин і органів у різні періоди життя організму, дослі-дженні реакції тканин на зовнішній вплив. Особливо широкі перспек-тиви перед практичною ембріологією відкриває розвиток досліджень щодо екології і захисту природи.
Як біологічна наука, ембріологія тісно пов'язана з анатомією, фізі-ологією, генетикою, біохімією, які є фундаментом ветеринарної освіти. Насамперед, ембріологія має важливе практичне значення у повсяк-денній роботі лікаря ветеринарної медицини, що пов'язане з заплід-ненням, вивченням патології вагітності, причин безпліддя тварин. Вона визначає вимоги до умов та шляхи щодо збереження здоров'я молодняка сільськогосподарських тварин, удосконалення порід і по-родоутворення.
Тісний зв'язок ембріології з птахівництвом, рибництвом, шовків-ництвом, що необхідно для створення сприятливих умов при штучній інкубації яєць сільськогосподарської птиці, розведенні риби, тутового шовкопряда.
Ембріологія сприяє формуванню загальноосвітнього розвитку та лікарського мислення.
2.2 Статеві клітини
Статеві клітини за своєю будовою подібні до соматичних клітин, але мають своєрідну будову містять гаплоїдний (половинний) набір хромосом.
Серед статевих клітин розрізняють два різновиди — сперматозоїди та яйцеклітини — овоцити.
Сперматозоїди (від лат зрегша - сімя, гооп — тварина). У різних видів сільськогосподарських та лабораторних тварин вони різняться своєю будовою. Однак для всіх них характерні морфологічні риси, що тісно пов'язані з їх основними біологічними особливостями, здатніс-
Розділ 2
Ембріологія
тю активно рухатися та запліднювати яйцеву клітину. При еякуляції сім'яна рідина містить мільярди сперміїв.
У сперматозоїда розрізняють голівку і хвостову частину Електрон-номікроскопічні дослідження показали, що голівка має невелике щіль-не ядро з гаплоїдним набором хромосом. У ядрах сперміїв містяться різні типи статевих хромосом, що несуть генетичну інформацію. По-ловина сперматозоїдів мають Х-хромосому, друга половина — У-хро-мосому Останні хромосоми належать до аутосом. Спермії, що несуть У-хромосому при злитті з яйцеклітиною започатковують організм самця; спермії — носії Х-хромосоми при злитті з яйцеклітиною запо-чатковують організм самки.
Передня частина головки має плоский чохлик, що вкриває акро-сому (від гр. акгоз - крайній, зота — тіло). Як чохлик, так і акросома є похідними комплексу Гольджі. В акросомі знаходяться ферменти — гі-алуронідаза і протеази, які здатні розчиняти оболонку яйцеклітини та стимулювати проникнення у неї спермія при заплідненні. Зовні головка спермія покрита плазмолемою, в ній містяться фібрили, які надають їй механічної міцності. Хвостова частина сперматозоїда нит-коподібна, вона складається із шийки, проміжної, основної та кінцевої частини (рис. 20).
За голівкою знаходиться шийка; в ній розміщена проксимальна центріоль і краніальна частина дистальної. Проксимальна центріоль під час запліднення переходить у зиготу і бере участь у її поділі. Голо-вна і кінцева частини хвоста сперматозоїда містить скоротливий апа-рат — аксонему; на її периферії розміщені дев'ять пар мікротрубочок і центральна пара. Навколо мікротрубочок по спіралі містяться міто-хондрії. Мітохондрії забезпечують енергією рухливість аксонеми. У складі проміжної частини містяться циркулярні пучки мікрофібрил. Головна частина хвостика утворена аксонемою і плазмолемою з тон-ким прошарком цитоплазми. В останній є значна кількість циркулярно орієнтованих тонофібрилярних структур. На межі між головною і про-міжною частинами хвоста навколо аксонеми знаходиться каудальна частина дистальної центріолі. Кінцева частина хвоста в своєму складі має осьову нитку з поодинокими скоротливими мікрофіламентами й покрита плазмолемою.
Біологічні властивості сперматозоїдів. В статевих шляхах самців (придаток сім'яника) у сперматозоїдів знижений обмін речовин, вони
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
майже нерухомі. В кінцевому відділі протоки додатка сперматозоїдів по-криваються тонкою ліпоїдно-білковою оболонкою, набувають однаково-го електричного заряду і починають відштовхуватися один від одного.
| Головка • |
| Шийка | |
| Хвіст і |
Середня частина
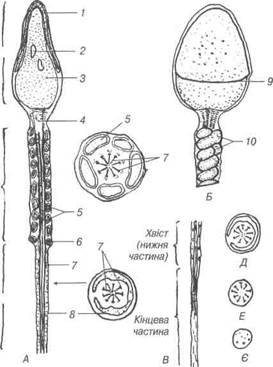
Рис. 20. Схема будови сперматозоїда самця:
1-головний чохлик; 2-вакуоль; 3-ядерний матеріал; 4-проксимальна центрі-оля; 5-мітохондрії; 6-каудальна частина дістальної центріолі; 7-осьові філа-менти; 8-оболонка; 9-край головиого чохлика; 10-мітохондріальна спіраль.
Сперматозоїди рухаються зі швидкістю 2-5 мм за хвилину. В ста-тевих шляхах птахів та ссавців вони рухаються прямолінійно, що за-лежить від симетрїї голівки. Спрямування руху сперматозоїдів визна-чається їх реотаксисом, тобто здатністю рухатися проти течії рідини. Реотаксис проявляється лише при порівняно слабкому зустрічному по-тоці слизу. В кислому середовищі голівки сперматозоїдів набрякають,
Розділ 2
Ембріологія
заокруглюються, втрачають асиметрію і тому вони набувають кругового руху, що заважає їм рухатися прямолінійно і усувається можливість зу-стрічі з яйцеклітинами. У сперматозоїда мало поживних речовин, тому він їх скоро втрачає і через 24-36 годин гине, якщо не зустрінеться з яй-цеклітиною.
Сперматозоїди досить чутливі до іонів двох- та трьохвалентних мета-лів, кислот, наркотиків, хініну. Глибоке охолодження до -78° або -196°С гальмує у них обмін речовин, що дає можливість продовжити термін їх зберігання на декілька років і використовувати тривалий час для заплід-нення.
Яйцеклітини — овоцити (від лат. оушп — яйце) — статеві кліти-ни самки.
За своєю будовою яйцеклітини відрізняються від соматичних клі-тин. Зрілі яйцеклітини мають гаплоїдний набір хромосом, після за-пліднення здатні розвиватися в новий організм. Вони містять більший чи менший запас поживних речовин (жовтка), необхідних для забез-печення зародкового розвитку організму, мають кортикальний шар ци-топлазми, обмежені спеціальними оболонками, які захищають зародок від шкідливої дії зовнішнього середовища. Для овоцитів характерна поляризована будова за рахунок нерівномірного розміщення поживних речовин.
Поживні речовини (жовток) мають вигляд гранул або складніших утворень, що мають вигляд шарів та платівок, утворених фосфоліпіда-ми, вуглеводами і протеїнами. Структурною одиницею жовтка є комп-лекс ліповітеліну і фосфовітину. Кожна платівка складається з щільнішої центральної і пухкої периферійної зони, зовні обмеженої осміофільною мембраною. Щільна зона утворена молекулами фосфовітина і має вигляд кристалічної сітки. Жовток синтезується при безпосередній участі ендо-плазматичної сітки та комплексу Гольджі.
Особливостями овоцитів є те, що вони містять значні запаси рибо-сом, ІРНК, тРНК, кількість яких в сотні разів перевищує їх в соматичних клітинах. В цитоплазмі яйцеклітини нагромаджуються запаси гістонів, тубуліну ліпофосфопротеїдів та жовтка.
Кортикальний шар ооплазми розміщений у периферійній части-ні овоцитів, він не має жовтка, містить більшість мітохондрій та гранул глікозаміногліканів. Кортикальний шар відіграє важливу роль в заплід-ненні (рис. 21).

| В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, ггстологія, ембріологія |
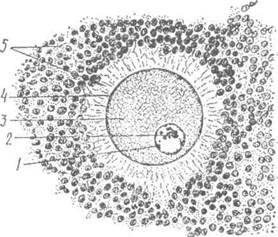
Рис. 21. Яйцеклітина ссавця:
1-ядро; 2-ядерце; 3-цитоплазма; 4-первинна оболонка; 5-вторинна оболонка (фолікулярні клітини).
\ / Яйцеклітини мають дві або три оболонки:,
^ 1) первинна оболонка (жовткова або відтелінова, це власне плаз-
молема);
2) вторинна оболонка - утворена фолікулярними клітинами та їх похідними. Останні формують блискучу (прозору) зону (від лат. гопа реііисісіа). Відростки фолікулярних клітин пронизують блискучу зону утворюючи променевий вінець (від лат. согопа гасііага). Вторинна оболонка виконує трофічну і захисну функції та запобігає поліспермії.
3) третинна оболонка - існує в яйцеклітинах плазунів, яйце-кладних, ссавців, та деяких риб (селахій) вона утворюється секретом яйцепроводів.
Для овоцитів характерна полярність, котра пов'язана з кількістю жовтка та розміщенням ядра. Та частина яйцеклітини, в якій накопи-чується жовток, є вегетативним полюсом, а протилежна, де знаходить-ся ядро і менше жовтка називається анімальним полюсом.
В залежності від кількості жовтка яйцеклітини поділяються на три групи: оліголецитальні (від гр. о1і§оз - мало, Іесігпоз — жовток), мезолецитальні (від гр. шезоз — середній) та полілецитальні (від гр.
Розділ 2
Ембріологія
роіі — багато). Залежно від розміщення жовтка яйцеклітини бувають телолецитальні (від гр. їеіоз — кінець) та ізолецитальні (від гр. ізоз — рівний).
Оліголецитальні овоцити характерні для плацентарних ссавців в зв'язку з внутрішньоутробним розвитком і живленням за рахунок мате-ринського організму. Винятком є овоцити представників примітивних ссавців (клоачні, частково сумчасті), які зберегли основні ознаки своїх попередників -- плазунів, у тому числі й телолецитальні овоцити. У те-лолецитальних овоцитів жовток концентрується на одному полюсі.
У сільськогосподарських тварин діаметр овоцита коливається від 100 до 140 мкм. Жовтка у них занадто мало, проте в ооплазмі речо-вини розподілені нерівномірно. На одному боці яйцеклітини нагрома-джуються РНП (рибонуклеопротеїди), тому на цій частині поверхні овоцита можна помітити серпоподібної форми скупчення базофільної речовини. Вважають, що цей базофільний серп відповідає і майбут-ній спині зародка. На майбутньому нейтральному боці його нагрома-джується жовток. Таким чином, у овоцитах ссавців ще до запліднення можна спостерігати полярність розміщення поживних речовин.
Овоцити ссавців містять органели загального призначення: ендо-плазматичну сітку, мітохондрії, комплекс Гольджі. На периферії цито-плазми знаходиться кортикальний шар, в якому знаходяться мітохон-дрії та гранули глікозаміногліканів. Включення ооплазми представле-ні жовтком у вигляді гранул, утворених фосфоліпідами, протеїнами та вуглеводами. Зовні від кортикального шару знаходиться плазмолема овоцита. Фолікулярні клітини формують і променевий вінець — сого-па гасііаіа - друга складова вторинної оболонки.
Мезолецитальні овоцити. Мають розмір до 2-3 мм, розміщені нерів-номірно. В ділянці анімального полюса мало жовтка, міститься ядро з великою кількістю ядерець. Більша частина жовтка розміщена з боку вегетативного полюса, на якому жовткові платівки щільно спаковані. Кортикальний шар товщиною 1 мкм містить гранули, а також чорний пігмент. Овоцити амфібії належать до телолецитального типу, чітко полярні, мають сполучнотканинну оболонку, зовні якої знаходиться слизовий шар, що являє собою третинну оболонку.
До полілецитального типу відносять яйцеклітини птахів. Власне яйцеклітиною птахів є жовток, розмір якого у курей досягає 3,5 см. В яйцепроводі яйцеклітина покривається третинними оболонками, до
5 - 8-305
В.ІІ. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
яких належать: зовнішній шар вітелінової мембрани, за її рахунок фор-муються градинки, які є зв'язками, на яких підвішений жовток, біл-кова оболонка, дві підшкаралупні оболонки, шкаралупа, побудована із оргапічмих волокон неорганічних речовин. Третинні оболонки вико-нують трофічну та захисну функції. Полілецитальні та телолецитальні яйцеклітини характерні для деяких риб, плазунів, птахів та яйценесу-чих ссавців (рис. 22).
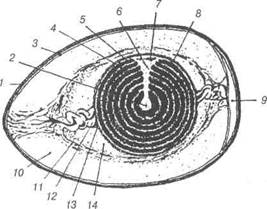
Рис. 22. Схема будови курячого яйця:
1-шкаралупа; 2-латебра; 3-підшкаралупна оболонка; 4-білий жовток; 5-жов-тий жовток; 6-ядро Пандера; 7-бластодерма; 8-жовткова оболонка; 9-пові-тряна камера; 10-зовнішній шар білка; 11-волокнистий шар білка; 12-халаза; 13-халазоутворюючий шар; 14-внутрішній шар білка.
2.3 Гаметогенез
Первинні статеві клітини — гаметобласти виникають у зародка на ранній стадії розвитку. В цей період вони ще не мають статевих осо-бливостей і являють собою великі клітини. Вони відрізняються від інших клітин зародка вмістом глікогену та активними ферментами, серед яких важливе місце належить лужній фосфатазі. За цією озна-кою їх виявляють, ставлячи відповідні гістохімічні реакції. Виникають ці клітини у птахів і ссавців в тимчасовому органі — мезодермі жовт-кового міхура, де формуються первинні кровоносні судини і де в цей період створюються сприятливі умови для живлення. Пізніше первин-
Розділ 2 Ембріологія
ні статеві клітини переносяться кров'ю або активно переміщуються в зародки гонад — сім'яники або яєчники, де і відбувається їх розвиток (рис.23).
Спосіб поділу статевих клітин, при якому дочірні клітини одер-жують вдвічі менше хромосом називається мейозом (від лат. шеіозіз — зменшення). Завдяки цьому поділу забезпечується постійність числа хромосом у наступних поколіннях. В результаті мейозу утворюються статеві клітини з одинарним (гаплоїдним) набором хромосом. Процес мейозу може тривати від кількох днів до кількох років.
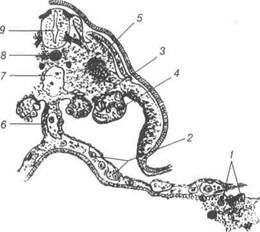
Рис. 23. Схема зрізу через середню частину тіла раннього зародка ссавця, що ілюструєвиникнення первинних статевих клітин у ентодермі жовткового міхура і їх міграцію до гонад:
1-первинні статеві клітини; 2-клітини, що мігрують до брижі; 3-клітини, які локалізуються у гонадах; 4-ембріональна нирка; 5-міотом; 6-дорзальна мезо-дерма; 7-аорта; 8-хорда; 9-нервова трубка.
Розвиток сперматозоїдів - сперматогенез.
Сперматогенез відбувається в звивистих сім'яних канальцях, у яких містяться сперматогенні клітини та клітини Сертолі (сустентоцити), останні утворюють мікросередовище для сперматогенних клітин.
В сперматогенезі розрізняють чотири періоди: розмноження, рос-ту, дозрівання, формування.
В.П. Новак, Ю.П. Бичков, М.Ю. Пшипенко Цитологія, гістологія, ембріологія
На стадії розмноження клітини називаються сперматогоніями. Вони межують з базальною мембраною, тобто займають периферій-ну частину звивистого сім'яного канальця. Сперматогонії невелико-го розміру містять диплоїдний набір хромосом. Серед них існує два різновиди: одні з них мають овальне світле ядро з розсіяним у ньо-му хроматином. Діляться вони мітозом, збільшуючись кількісно. Це стовбурові клітини. Сперматогонії другого різновиду мають кулясте ядро, що містить зернистий хроматин. Такі сперматогонії перестають ділитися і переходять в стадію росту, перетворюючись у спермато-цити першого порядку. Вони збільшені в розмірі, зміщені в напрямі просвіту звивистого сім'яного канальця, зберігають диплоїдний на-бір хромосом, проходять профазу мейозу. Перед початком профази у сперматоциті першого порядку подвоюється ДНК, а також відбу-ваються складні зміни в перерозподілі спадкового матеріалу в ядрі, в зв'язку з цим спостерігаються такі фази: лептонеми, зигонеми, па-хінеми, диплонеми.
В стадії лептонеми (від гр. Іерїоз - тонкий, петае — нитка) помітна ядерна мембрана, ядерце та хромосоми, що мають вигляд тонких ниток.
В зигонемі (від гр. 2І§оІІ8 -з'єднання) — гомологічні хромосоми розміщуються парами, кон'югуючись за довжиною і утворюють біва-ленти, або діади. При цьому гомологічні хромосоми обмінюються ге-нами, що забезпечує мінливість спадкового матеріалу в ряді поколінь.
На стадії пахінеми (від гр. распуз — твердий) обидві аутосоми спі-ралізуються, потовщуються, стають коротшими, залишаються у тіс-ному контакті по всій довжині. На цій стадії закінчується кон'югація хромосом.
В диплонемі (від гр. сііріоз — подвійний) в кожній із кон'югую-чих гомологів з'являються по парі сестринських хроматид (рис. 24). Хроматиди пов'язані центромерою, що перетворює біваленти в тетра-ди. В кожній парі виникає поздовжня щілина, і у двох кон'югованих хромосомах утворюється чотири хроматиди. В тетрадах з'являються перехрести хромосом (хіазми), що свідчить про обмін гомологічними частинами між несестринськими хроматидами в тетраді. В стадії ди-плонеми зникають сили гомологічного притягання і виникають сили відштовхування між сестринськими хроматидами. Після цього хро-мосоми ще більше спіралізуються, потовщуються і відокремлюються одна від другої.
| Розділ 2 Ембріодогія |
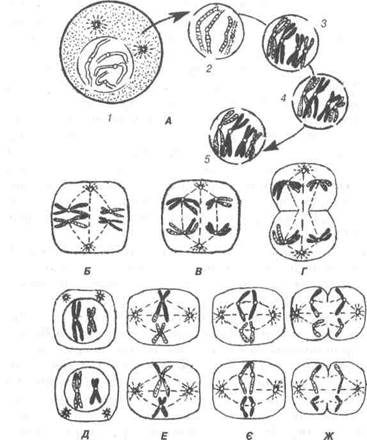
Рис. 24. Схематичне зображення основних стадій мейозу (пояснення в тексті):
А — перше ділення мейозу; 1-лептотена; 2-зиготена; 3-пахітена; 4-диплотена; 5-діакінез.
За періодом росту сперматоцити першого порядку вступають в пе-ріод дозрівання, який відбувається у два етапи. Після першого — ре-дукційного поділу сперматоцита першого порядку утворюється два сперматоцити другого порядку. Кожний з них одержує половину від кількості хромосом сперматоцита першого порядку. Сперматоцити
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
другого порядку містять диплоїдний набір хромосом. Кожна із хромо-сом має по дві хроматиди. Сперматоцити другого порядку менших розмірів. Перед другим поділом дозрівання у сперматоциті другого по-рядку не синтезується ДНК і не подвоюється хромосомний матеріал.
У результаті другого — екваційного поділу дозрівання утворю-ються сперматиди — клітини з гаплоїдним набором хромосом, кожна з яких складається з однієї хроматиди. При цьому пари хроматид, які з'єднані між собою центромерою, роз'єднуються і розподіляються між сперматидами, стають хромосомами. Сперматиди малого розміру, бід-ні на хроматин, містять мітохондрії, центросому, комплекс Гольджі та інші органели. Залежно від стадії розвитку сперматиди можуть мати округлу чи витягнуту форму. Сперматиди округлої форми розміщу-ються біля просвіту звивистого сім'яного канальця, а витягнутої фор-ми — занурюються в цитоплазму клітин Сертолі, в якій вони перехо-дять в період формування.
Таким чином, у фазі дозрівання в результаті мейозу із кожного сперматоцита першого порядку виникає чотири сперматиди з одинар-ною кількістю хромосом. Статеві X і У-хромосоми рівномірно розподі-ляються між сперматидами.
Стадія формування відбувається шляхом перетворення сперма-тид в сперматозоїди. При цьому їх ядра ущільнюються, набувають овальної форми, перетворюються у голівки сперматозоїдів і займають ексцентричне положення в клітині. Одночасно вони частково занурю-ються в цитоплазму клітин Сертолі, що зумовлює перетворення спер-матид в сперматозоїди. Вважається, що клітини Сертолі забезпечують трофіку та фагоцитують залишки, що виникають в процесі дегенерації неповноцінних клітин. Деталі перетворення сперматиди в спермато-зоїда можна виявити за допомогою електронної мікроскопії. Так, час-тина цитоплазми сперматиди, що містить апарат Гольджі, концентру-ється на апікальному кінці голівки сперматозоїда. У цій зоні виникає ущільнена гранула — акробласт. Він збільшується в розмірі і у вигляді чохлика покриває ядро. В середині акробласта утворюється акросома. Центросома входить до складу шийки, вона складається із двох цен-тріолей, зміщується в протилежний кінець сперматиди. Проксимальна центріоля прилягає до поверхні ядра, а дистальна поділяється на дві частини. Від краніальної частини дистальної центріолі помітно поча-ток формування джгутика сперматозоїда, каудальна частина центріолі
Розділ 2
Ембріологія
набуває вигляд кільця, зміщуючись по джгутику, це кільце визначає задню межу середньої частини сперматозоїда.
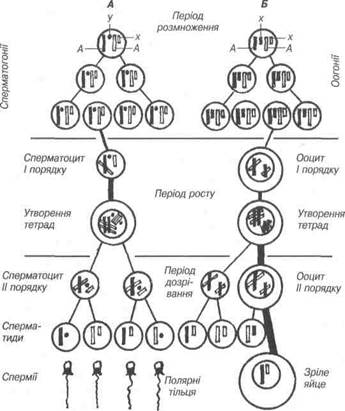
Рис. 25. Схема сперматогенезу (А) і овогенезу (Б).
Батьківські хромосоми чорні, материнські світлі. А — аутосоми; Х,У — стате-ві хромосоми; в періоді розмноження показані три покоління сперматогоній і оогонії.
Під час росту хвоста цитоплазма сповзає з ядра і концентрується у проміжній частині. Мітохондрії розміщуються спірально між про-ксимальною центріолею та кільцем. Цитоплазма сперматиди під час її перетворення в сперматозоїд значно редукується. В ділянці голівки
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
вона зберігається лише у вигляді тонкого шару, що покриває акросому. Незначна її кількість сповзає в дистальному напрямку і утворює тонку оболонку хвостика, а його кінцева частина зовсім не має цитоплазми. Невикористана частина цитоплазми відокремлюється і розпадається у просвіті звивистого сім'яного канальця.
Таким чином, кожний початковий сперматогоній дає початок чо-тирьом сперматозоїдам. Сперматогенний епітелій занадто чутливий до впливу інтоксикації, авітамінозу, дії іонізуючого випромінювання. При недостатньому живленні сперматогенез послаблюється або зовсім зникає, а сперматогенний епітелій атрофується.
Овогенез — розвиток статевих клітин самки.
На відміну від сперматогенезу, овогенез у тварин різних класів зна-чно різноманітніший і залежить від біології розмноження. Він перебі-гає протягом трьох періодів: розмноження, росту та дозрівання.
Період розмноження. Якщо розмноження сперматогоній почина-ється після досягнення статевої зрілості і продовжується протягом усієї статевої активності самця, то в овогенезі період розмноження від-бувається лише у зародка і закінчується після народження плода. Піс-ля народження кількість статевих клітин не збільшується. Вони мають малий розмір, їх називають овогоніями. Разом з покриваючими їх фо-лікулярними клітинами називаються примордіальними фолікулами.
Період росту. В період росту клітини називаються овоцитами першого порядку 3 самого початку свого існування овоцит першого порядку вступає в профазу мейозу. В цей період, як і в сперматоци-тах першого порядку, послідовно відбуваються такі стадії: лептонема, зигонема, пахінема й диплонема. Цей період у тварин різних класів характеризується значною повільністю. Так, він може відбуватися за кілька днів, тижнів і навіть десятків років. Останнє спостерігається у ссавців, де розмноження овогоній закінчується іще до народження плода, а овуляція настає не раніше статевої зрілості. Після диплоне-ми хромосоми розходяться, деспіралізуються, стають непомітними до кінця періоду росту. Овоцит першого порядку інтенсивно синтезує ре-човини за допомогою хромосом.
Нагромадження поживних речовин овоцитом першого порядку поділяють на превітелогенез (фаза невеликого росту) та вітелогенез (фаза великого росту). Превітелогенез характеризується ростом цито-плазми і переважним синтезом інформаційної РНК, яка пізніше буде
Розділ 2
Ембріологія
матрицею для синтезу білків. Такий інтенсивний синтез у овогенезі ін-формаційної РНК пов'язаний з тим, що він іде на запас. Не більше тре-тини його використовується безпосередньо в овогенезі, тоді як основна частина білкового синтезу починається після запліднення. У цей час в цитоплазмі утворюються органели, необхідні для синтезу: гранулярна цитоплазматична сітка, мітохондрії, комплекс Гольджі.
У фазі великого росту синтезуються запасні поживні речовини, що нагромаджуються у цитоплазмі і використовуються лише після заплід-нення. Основні з них жовток, жир, глікогени. Жовток являє собою крис-талізовані речовини білкової природи у формі фосфопротеїнів. Криста-ли мають вигляд гранул чи платівок різної форми. Кількість жовтка, що нагромаджується в яйцевій клітині, неоднакова у тварин різних груп. В полілецитальних яйцеклітинах, наприклад курей, жовток відкладається інтенсивно. Так, за добу відкладається шар жовтка товщиною до 2,5 мм. У курей перед овуляцією яйцеклітина досягає 35 мм (Б.Карлсон, 1983). Яйцеклітини ссавців значно менші за розміром, їх ріст може продовжу-ватися роками, наприклад у корів — 10 років і більше.
В синтезі жовтка в цей час бере участь весь організм, особливо печінка. Запас поживних речовин у вигляді жовтка називають дейто-плазмою. В процесі нагромадження жовтка майже завжди виявляють полярність овоцита. Полюс, багатий на жовток, називають вегетатив-ним, протилежний, куди зміщується ядро — анімальним. У фазі вели-кого росту органели розсіюються по цитоплазмі з подальшою концен-трацією на периферії.
Період дозрівання відбувається після проникнення в яйцеву клі-тину сперматозоїда (у аскариди), у ланцетника спермій проникає піс-ля першого поділу; у більшості тварин він проникає в яйце лише після дозрівання, яке настає після виходу яйцеклітини із яєчника.
Дозрівання овоцита — це складні перетворення у ядрі. Як і при спер-матогенезі, воно складається із двох послідовних поділів, які надають яйцеклітині гаплоїдного стану. Однак, в дозріванні яйцеклітини, на від-міну від сперматогенезу, один первинний овоцит утворює лише одну зрілу яйцеклітину, яка повністю зберігає жовток, що утворився в період росту, і три неповноцінні клітини — полярні (редукційні) тільця. Вони містять лише ядро без цитоплазми. При цьому ядро первинного овоцита ділить-ся порівну між двома дочірніми клітинами, тобто між овоцитом другого порядку і першим редукційним тільцем. Подальший поділ приводить до
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
того, що овоцит другого порядку також ділиться нерівномірно — вини-кає велика зріла яйцеклітина та друге редукційне тільце. В подальшому редукційні тільця гинуть. їх призначення полягає у тому, що дозріваюче за їх допомогою яйце звільняється від половини хромосом. Під час дру-гого поділу дозрівання яйцеклітина втрачає свою центросому.
Дата добавления: 2014-12-18; просмотров: 5518;