Электрический синапс
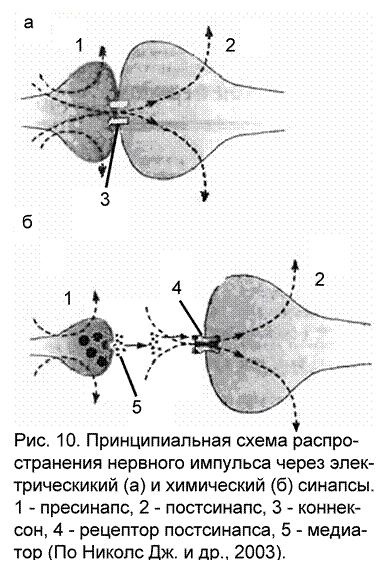 Существование электрических синапсов в природе подозревалось давно, но описаны они были только в 1957-59 гг. Такой тип клеточных контактов более характерен для беспозвоночных животных, у позвоночных он встречается реже. Отличительной чертой электрических синапсов является очень узкая синаптическая щель, а электрические взаимодействия возможны благодаря высокопроводящим (низкоомным) соединениям – щелевым контактам (рис. 10а). Щелевым контактом называется участок мембран двух клеток, в каждой из которых имеется скопление особых частиц, именуемых коннексонами.Они состоит из шести белковых субъединиц, образующих круг с внешним диаметром около 10 нм и внутренним диаметром 2 нм. На сопряженной клетке находится точно такая же структура. Совместно они пронизывают зазор между мембранами (2-3 нм). Внутри коннексонов имеется полость, через которую происходит перемещение ионов и мелких молекул между клетками. При распространении нервного импульса потери виде утечки тока через синаптическую щель в окружающую среду минимальны. Именно эта особенность позволила распознать в эксперименте электрические синапсы. Впервые это обнаружили Эд. Фершпен и Д. Поттер при исследовании синапсов речного рака. В стволе мозга млекопитающих также найдены области с электрическими синапсами: в среднем мозге, между телами клеток ядра тройничного нерва, в вестибулярном ядре Дейтерса между телами клеток и окончаниями аксонов, в продолговатом мозге между дендритами. Есть данные о присутствии электрических синапсов в колонках коры больших полушарий. Электрические синапсы позволяют синхронизировать активность групп нейронов, что дает возможность получать постоянные стереотипные реакции организма при многократных воздействиях.
Существование электрических синапсов в природе подозревалось давно, но описаны они были только в 1957-59 гг. Такой тип клеточных контактов более характерен для беспозвоночных животных, у позвоночных он встречается реже. Отличительной чертой электрических синапсов является очень узкая синаптическая щель, а электрические взаимодействия возможны благодаря высокопроводящим (низкоомным) соединениям – щелевым контактам (рис. 10а). Щелевым контактом называется участок мембран двух клеток, в каждой из которых имеется скопление особых частиц, именуемых коннексонами.Они состоит из шести белковых субъединиц, образующих круг с внешним диаметром около 10 нм и внутренним диаметром 2 нм. На сопряженной клетке находится точно такая же структура. Совместно они пронизывают зазор между мембранами (2-3 нм). Внутри коннексонов имеется полость, через которую происходит перемещение ионов и мелких молекул между клетками. При распространении нервного импульса потери виде утечки тока через синаптическую щель в окружающую среду минимальны. Именно эта особенность позволила распознать в эксперименте электрические синапсы. Впервые это обнаружили Эд. Фершпен и Д. Поттер при исследовании синапсов речного рака. В стволе мозга млекопитающих также найдены области с электрическими синапсами: в среднем мозге, между телами клеток ядра тройничного нерва, в вестибулярном ядре Дейтерса между телами клеток и окончаниями аксонов, в продолговатом мозге между дендритами. Есть данные о присутствии электрических синапсов в колонках коры больших полушарий. Электрические синапсы позволяют синхронизировать активность групп нейронов, что дает возможность получать постоянные стереотипные реакции организма при многократных воздействиях.
Химический синапс
В 1892 году Лэнгли предположил, что синаптическая передача в вегетативных ганглиях млекопитающих имеет химическую природу, а не электрическую. Примерно через 10 лет Эллиот обнаружил, что экстракт, полученный из надпочечников, — адреналин — производит на клетки такое же воздействие, как и стимуляция симпатических нервов. Это позволило предположить, что адреналин может секретироваться нейронами, и выделятся нервными окончаниями при проведении возбуждения. Однако лишь в 1921 году Леви поставил опыт, в котором была установлена химическая природа передачи в вегетативных синапсах между блуждающим нервом и сердцем. Он наполнял физиологическим раствором сосуды сердца лягушки и стимулировал блуждающий нерв, вызывая замедление сердцебиений. Когда жидкость из заторможенного стимуляцией сердца была перенесена на второе нестимулированное сердце, оно в результате такого воздействия также начинало биться медленнее. Было очевидно, что стимуляция блуждающего нерва вызывала освобождение в раствор тормозящего вещества. В последующих экспериментах Леви и его коллеги показали, что ацетилхолин полностью воспроизводил эффекты этого вещества.
Удивительно, что идея этого эксперимента пришла Леви во сне, он записал ее, но поутру не смог разобраться в том, что написал ночью. К счастью, сон повторился, и в этот раз Леви не стал рисковать: он помчался в лабораторию и поставил этот эксперимент. Вот как он вспоминает эту ночь: По серьезному размышлению, в холодных лучах утреннего света, я бы никогда не поставил этого эксперимента. То, что блуждающий нерв может освобождать тормозящее вещество, было очень маловероятно; еще менее вероятным было то, что химическое вещество, которое предположительно должно было действовать на коротком расстоянии между нервным окончанием и мышцей, освобождалось бы в таком большом количестве, что сохраняло бы эффективность после разведения в перфузирующей жидкости.
В начале 1930-х годов роль ацетилхолина в синаптической передаче в ганглиях вегетативной нервной системы была окончательно установлена Фельдбергом и его сотрудниками. Дейл, британский физиолог и фармаколог, установил роль ацетилхолина при передаче возбуждения в нервно--мышечном синапсе.
Принципиальным отличием химического синапса передача раздражения с пресинапса на постсинапс при помощи биологически активных веществ, называемых медиаторами (трансмиттерами). В связи с этим появляются и отличия в морфологии химического синапса (рис. 10б; приложение 2):
 пресинаптическая область обязательно содержит везикулы (замкнутые в собственные мембраны полости), наполненные медиатором. Везикулы способны к перемещениям, когда они подходят к мембране нервного окончания, то их мембраны сливаются, а медиатор высыпается в синаптическую щель.
пресинаптическая область обязательно содержит везикулы (замкнутые в собственные мембраны полости), наполненные медиатором. Везикулы способны к перемещениям, когда они подходят к мембране нервного окончания, то их мембраны сливаются, а медиатор высыпается в синаптическую щель.
синаптическая щель имеется между пре- и постсинаптическими мембранами. Величина щели неодинакова в различных синапсах. Так, в приведенном ранее примере нервно-мышечного окончания, где медиатором является ацетилхолин, ширина синаптической щели достигает 30 нм. Пространство синаптической щели заполнено межклеточной жидкостью, а в нервно-мышечном окончании коллоидом (поэтому часто синаптическую щель в нервно-мышечном окончании рассматривают как базальную мембрану).
постсинаптическая мембрана содержит на своей поверхности рецепторы – белки, обладающие большим сродством к медиатору. Определенному типу медиатора соответствует определенный тип рецептора, например, в нервно-мышечном окончании – холинорецепторы, они распознают медиатор в окружающей среде и вступают с ним во взаимодействие. В результате этого взаимодействия происходят структурные изменения на постсинаптической мембране, в результате которых изменяется ионная проводимость через постсинапс. Это, в свою очередь, является причиной дальнейшего распространения нервного импульса (в случае, если медиатор оказывает тормозящий эффект, то развитию процессов торможения на постсинаптической мембране).
Кроме ацетилхолина в качестве медиатора могут выступать также:
аминокислоты: глутамат, глицин, аспартат
биогенные амины: дофамин, g-аминомасляная кислота (ГАМК), норадреналин, адреналин, серотонин, гистамин
производные пурина: АТФ, АДФ, АМФ, аденозин
пептиды: энкефалины, эндорфины, вещество Р и многие другие.
Синапсы и нейроны, в соответствии с видом выделяемого медиатора называются по типу медиатора с добавлением окончания –эргический: холинэргический, адренэргический, ГАМКэргический, гистаминэргический и т.п.
Химические синапсы наиболее распространены в ЦНС позвоночных. В настоящее время известно, что нейрон может синтезировать и выделять несколько медиаторов (сосуществующие медиаторы). Нейроны также обладают нейромедиаторной пластичностью – способностью менять основной медиатор в процессе развития.
Рефлекторная дуга
При помощи синаптических контактов нейроны соединяются в нейронные цепи, которые в свою очередь являются структурной основой рефлекторной деятельности организма. По своему положению в цепи и в соответствии с выполняемой работой, нейроны подразделяются на чувствительные (афферентные), вставочные (промежуточные, центральные) и эффекторные (эфферентные) (рис. 11).
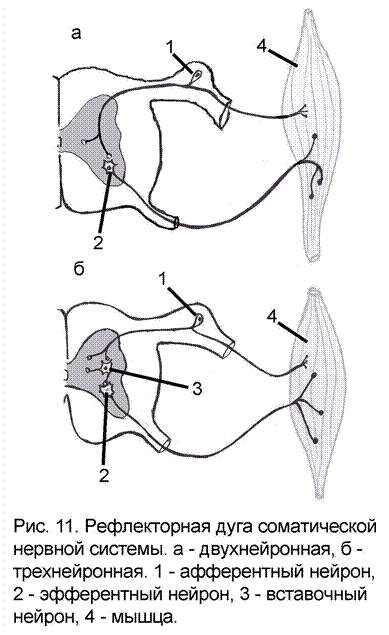 Перечисленные нейроны образуют рефлекторную дугу - путь, по которому проходят нервные импульсы от рецептора к исполнительному органу при осуществлении рефлекса. В ответ на рефлекторное воздействие от рабочего органа в ЦНС также поступают нервные импульсы, которые несут информацию о происходящих изменениях на периферии. Это звено работает по принципу обратной связи, осуществляя саморегуляцию деятельности организма.
Перечисленные нейроны образуют рефлекторную дугу - путь, по которому проходят нервные импульсы от рецептора к исполнительному органу при осуществлении рефлекса. В ответ на рефлекторное воздействие от рабочего органа в ЦНС также поступают нервные импульсы, которые несут информацию о происходящих изменениях на периферии. Это звено работает по принципу обратной связи, осуществляя саморегуляцию деятельности организма.
Принимая во внимание сигналы, поступающие от рабочего органа в ЦНС в ответ на рефлекторное воздействие, можно сказать, что рефлекторная дуга представляет собой сложный структурно-функциональный комплекс, замкнутый в кольцо. Компонентами такого рефлекторного комплекса являются:
рецептор – специализированная клетка, воспринимающая раздражение. Эта клетка переводит специфическую энергию раздражителя (механическую, термическую, химическую) в энергию нервного импульса.
афферентный путь – отрецептора к ЦНС
рефлекторный центр – совокупность нейронов, лежащих в ЦНС, в которых происходит обработка информации и формируется ответная реакция (образован вставочными нейронами)
эфферентный путь – от ЦНС на периферию
рабочий орган – мышца, железа
обратная связь – из органов, деятельность которых изменяется под влиянием эфферентных импульсов, посылаемых мозговыми центрами, обратно в мозг поступает информация о характере этих изменений (т.н. обратная афферентация). Различают два вида обратной связи: положительную – вызывает усиление ответной реакции, и отрицательную – вызывает торможение ответной реакции.
В зависимости от сложности строения рефлекторные дуги делят на простые и сложные.
Простые состоят из двух нейронов – чувствительного и двигательного, например, рефлекторная дуга коленного рефлекса. Теоретически в такой дуге может быть только один синапс, по другому она еще называется моносинаптической (приставка моно- означает «один»). На практике соответственно регистрируемому времени рефлекса, оказывается, что даже двухнейронная рефлекторная дуга имеет от 3 до 5 синапсов т.е. является полисинаптической (приставка поли- означает «много»). Простых рефлекторных дуг в организме человека немного, не более двух процентов от общего количества.
Сложные рефлекторные дуги содержат три и более нейронов – чувствительный, вставочный (от одного до десятков и сотен) и двигательный. Именно такие рефлекторные дуги наиболее распространены, а 90% нейронов ЦНС относятся к категории вставочных (приложение 3).
Также рефлекторные дуги, а, следовательно, и сами рефлексы можно классифицировать по следующим признакам:
в зависимости от того, где расположены рецептивные поля
o экстероцептивные – имеют рецепторы на внешней поверхности тела
o висцеро- или интероцептивные – имеют рецепторы во внутренних органах
o проприоцептивные – имеют рецепторы, регистрирующие движения (от мышц, связок, сухожилий)
по характеру ответной реакции
o соматические – проявляются в изменении функции с участием соматической нервной системы – произвольные движения.
o вегетативные – вызывают изменения в работе висцеральных органов.
в зависимости от того, где располагается рефлекторный центр данной дуги:
o спинальные – замыкаются на уровне спинного мозга
o бульбарные – на уровне продолговатого мозга
o мезэнцефальные – на уровне среднего мозга
o диэнцефальные – на уровне промежуточного мозга
o корковые – коры больших полушарий.
Дата добавления: 2019-07-26; просмотров: 1042;