Морфологические особенности глаза человека
Орган зрения состоит из глазного яблока (глаза), зрительного нерва и вспомогательных органов глаза. Глаз состоит из оптической и фоторецепторной частей и имеет три оболочки: белочную, сосудистую и сетчатую. Оптическая система глаза, обеспечивающая его светопреломляющую функцию, состоит из роговицы, передней и задней камер глаза, зрачка, хрусталика и стекловидного тела.
Глазное яблоко имеет шаровидную форму; его объем составляет 7,45 см3. В нем выделяют передний и задний полюсы. Передний полюс - это наиболее выступающая точка роговицы, задний полюс расположен латерально от места выхода зрительного нерва. Соединяющая оба полюса условная линия - это наружная ось глаза. Она равна примерно 24 мм и находится в плоскости меридиана глазного яблока. Плоскость, перпендикулярная наружной оси глаза, разделяет глазное яблоко на две половины - переднюю и заднюю и образует экватор, равный 23,3 мм.
Глазное яблоко состоит из ядра (хрусталик, стекловидное тело), покрытого тремя оболочками: наружной (фиброзной), средней (сосудистой) и внутренней (сетчатой) (рис. ).
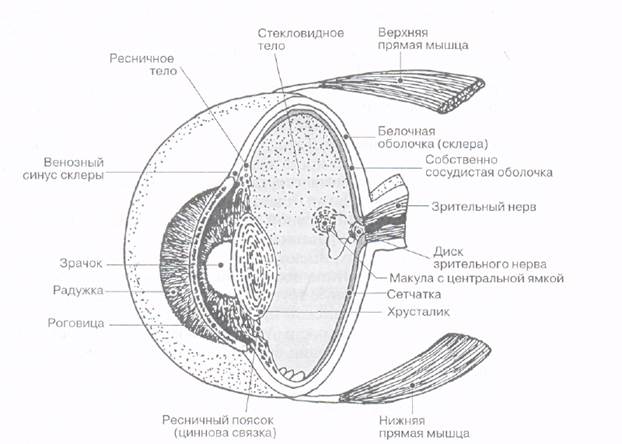

Наружная (фиброзная, или белочная) оболочка глазного яблока подразделяется на задний отдел - склеру и прозрачный передний - роговицу.
Склера представляет собой плотную соединительнотканную оболочку, которая в задней части имеет толщину 0,3-0,4 мм, а в близи роговицы - 0,6 мм. Склера образована пучками коллагеновых волокон, между которыми залегают уплощенные фибробласты с небольшим количеством эластических волокон. Сзади на склере находится решетчатая пластинка, через которую проходят волокна зрительного нерва. В зоне соединения склеры с роговицей имеется венозный синус склеры, или шлеммов канал, через который жидкость из передней камеры глаза оттекает в венозную сеть.
Роговица (cornea) - это прозрачная выпуклая пластинка блюдцеобразной формы (она напоминает слегка выпуклое часовое стекло). В отличие от склеры она лишена кровеносных сосудов (ее питание происходит за счет диффузии жидкости из сосудов лимба и передней камеры глаза) и совершенно прозрачна. Роговица состоит из пяти слоев: переднего эпителия, передней пограничной пластинки (боуменовой мембраны), собственного вещества, задней пограничной пластинки (десцеметовой оболочки) и заднего эпителия (эндотелия роговицы). Первый и третий слои богаты нервными окончаниями.
Сосудистая оболочка глазного яблока расположена под склерой. Ее толщина достигает всего 0,1-0,2 мм. Она богата кровеносными сосудами и состоит из трех частей: собственно сосудистой оболочки (choroidea), ресничного тела (corpus ciliare, цилиарное тело) и радужки (iris).
Собственно сосудистая оболочка - это густая сеть переплетающихся между собой артерий и вен. Между ними располагается рыхлая волокнистая соединительная ткань, богатая крупными пигментными клетками.
Впереди сосудистая оболочка переходит в утолщенное ресничное (цилиарное) тело кольцевидной формы, которое участвует в аккомодации глаза, поддерживая, фиксируя и растягивая хрусталик. Ресничное тело делится на две части: внутреннюю - ресничный венец, и наружную - ресничный кружок. От поверхности ресничного кружка по направлению к хрусталику отходят 70-75 ресничных отростков длиной около 2 мм каждый. К этим отросткам прикрепляются волокна ресничного пояска (цинновой связки), идущие к хрусталику. Из сосудов ресничных отростков (в области ресничного венца) выделяется водянистая влага, заполняющая камеры глаза; эпителиальные клетки, покрывающие ресничное тело и его отростки, принимают участие в ее образовании. Сзади ресничное тело и его отростки покрыты ресничной частью сетчатки.
Большая часть ресничного тела - это ресничная, или цилиарная, мышца. Она образована продольными, циркулярными и радиальными пучками миоцитов. Сокращение мышцы приводит к расслаблению волокон ресничного пояска (цинновой связки), хрусталик расправляется, округляется, вследствие этого выпуклость хрусталика и его преломляющая сила увеличивается, происходит аккомодация на близлежащие предметы. В старческом возрасте миоциты цилиарной мышцы частично атрофируются, что приводит к нарушению аккомодации.
Ресничное тело кпереди продолжается в радужку, которая представляет собой круглый диск с отверстием в центре –зрачок (pupilla). Радужка расположена между роговицей и хрусталиком. Она отделяет переднюю камеру (ограниченную спереди роговицей) глаза от задней (ограниченной сзади хрусталиком). Радужка состоит из пяти слоев: переднего (эпителий, являющийся продолжением эпителия, покрывающего заднюю поверхность роговицы), наружного пограничного слоя, сосудистого слоя, внутреннего пограничного слоя и пигментного слоя. В толще сосудистого слоя проходят две мышцы - сфинктер, или суживатель, зрачка (циркулярно ориентированные мышечные пучки), который иннервируется постганглионарными парасимпатическими волокнами клеток ресничного узла, и расширитель, или дилататор зрачка (мышечные пучки имеют радиальное направление), иннервируемый постганглионарными симпатическими волокнами клеток, лежащих в верхнем шейном узле. Пигментный слой радужной оболочки является продолжением эпителия, покрывающего цилиарное тело и реснитчатые отростки; он двухслойный. Различное количество и качество пигмента меланина обусловливает цвет глаз - карий, черный (при наличии большого количества пигмента) или голубой, зеленоватый- если мало пигмента (цвет глаз у крупного рогатого скота темно-коричневый, у лошади - темно- и светло-коричневый, у овцы - желто-бурый, у козы - голубоватый или желтоватый, у собаки - от темно-коричневого до желтого, иногда голубого цвета, у кошки - желтый или зеленый, иногда голубой; у пегих животных глаза часто бывают разного цвета). Лишь у альбиносов, в организме которых пигмент вообще отсутствует, радужная оболочка не окрашена, сквозь нее просвечивают кровеносные сосуды и поэтому радужка кажется красной.
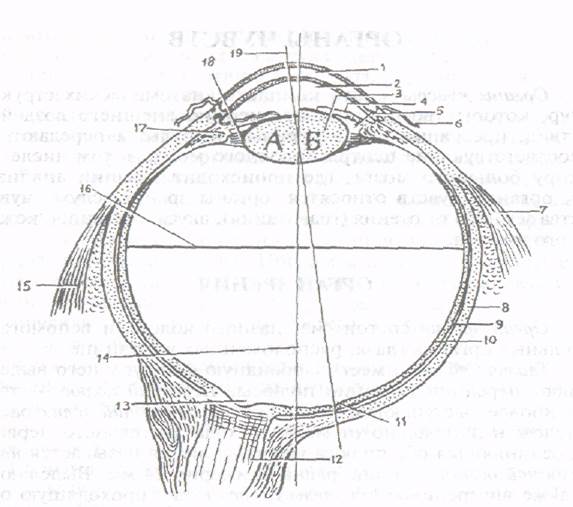
Рис. . Глазное яблоко. Разрез в горизонтальной плоскости (схема): Хрусталик при расслаблении (а) и сокращении (б) ресничной мышцы.
1 - роговица, 2 - передняя камера глаза, 3 - хрусталик, 4 - радужка, 5 - задняя камера глаза, 6 - конъюнктива, 7 - латеральная прямая мышца, 8 - белочная оболочка (склера), 9 - собственно сосудистая оболочка, 10 - сетчатка, 11 - центральная ямка, 12 - зрительный нерв, 13 - диск зрительного нерва (слепое пятно), 14 - наружная ось глаза, 15 - медиальная прямая мышца, 16 - поперечная ось глаза, 17 - ресничное тело, 18 - ресничный поясок, 19 - зрительная ось глаза
Внутренняя (светочувствительная) оболочка глазного яблока - сетчатка (retina) - на всем протяжении прилежит изнутри к сосудистой оболочке. Сетчатка состоит из двух листков: внутреннего - светочувствительного (нервная часть) и наружного - пигментного. Сетчатка делится на две части - заднюю зрительную и переднюю (ресничную и радужковую), которая не содержат фоторецепторов. Место выхода из сетчатки зрительного нерва называется диском зрительного нерва, или слепым пятном - здесь также отсутствуют фоторецепторы. В его наличии можно убедиться при рассматривании задней стенки глазного яблока (глазного дна) при помощи вогнутого зеркала- офтальмоскопа. При этом виден бледно окрашенный участок, от которого расходятся кровеносные сосуды. В центре диска в сетчатку входит центральная артерия сетчатки. Со всей сетчатки к слепому пятну сходятся нервные волокна, образующие зрительный нерв. В существовании слепого пятна можно также убедиться с помощью опыта Мариотта (просмотр с расстояния 10-25 см рисунка, содержащего на темном фоне белый крест и белый круг), а также при проведении кампиметрии.
На расстоянии около 4 мм от зрительного диска (по направлению к наружному краю глаза) имеется желтое пятно, диаметр которого составляет примерно 1,5 мм. Желтая окраска обусловлена присутствием каротиноидов. В области желтого пятна отсутствуют кровеносные сосуды. В его центре находится так называемая центральная ямка, которая содержит только колбочки. Она является местом наилучшего видения глаза. По мере удаления от центральной ямки количество палочек возрастает, а число колбочек убывает.
В сетчатке различают 10 слоев. Однако с точки зрения функции главной является радиально ориентированная трехнейронная цепь, состоящая из наружного - фоторецепторного (палочек и колбочек), среднего - ассоциативного, и внутреннего - ганглионарного, компонентов. От каждой фоторецепторной клетки отходит тонкий отросток, заканчивающийся в наружном сетчатом слое утолщением, которое образует синапс с отростками биполярных клеток (они располагаются во внутреннем зернистом слое сетчатки). В свою очередь биполярные нейроны, выполняющие функцию ассоциативных нейронов, передают сигнал крупным оптикоганглионарным невроцитам (ганглиозные клетки, которые лежат в ганглионарном слое сетчатки). Их аксоны и образуют зрительный нерв. В сетчатке также располагается горизонтальные и амакриновые клетки, выполняющие ассоциативную функцию, и элементы нейроглии - радиальные глиоциты (мюллеровы клетки), астроциты и микроглиальные клетки. Мюллеровые клетки протягиваются почти через всю толщину сетчатки перпендикулярно ее слоям. Они занимают практически все пространства между нейронами сетчатки и их отростками. Своими основаниями мюллеровы клетки формируют внутреннюю глиальную пограничную мембрану, которая отделяет сетчатку от стекловидного тела. Апикальные отростки мюллеровых клеток формируют у основания фоторецепторов наружную глиальную пограничную мембрану. Многочисленные отростки мюллеровых клеток оплетают тела нейронов в области синаптических связей, выполняя поддерживающую и трофическую функции. Вместе с астроцитами, которые расположены, в основном, во внутренних слоях сетчатки, они окружают капилляры, образуя гемато-ретинальный барьер. Немногочисленные клетки микроглии располагаются во всех слоях сетчатки, выполняя фагоцитарную функцию.
Васкуляризация (кровоснабжение) сетчатки осуществляется центральной артерией сетчатки, которая вместе со зрительным нервом проникает в нее в области зрительного диска и разделяется на расходящиеся ветви. Сначала эти ветви располагаются между стекловидным телом и внутренней глиальной пограничной мембраной. Затем они проникают в слои сетчатки, образуя капиллярное скопление, которое доходит до внутреннего ядерного слоя (то есть до слоя, в котором находятся ядра биполярных, амакринных, горизонтальных и мюллеровых клеток). Расположенные кнаружи от него слои сетчатки получают питание диффузно из хориокапиллярного слоя сосудистой оболочки через слой пигментного эпителия (рис. ).
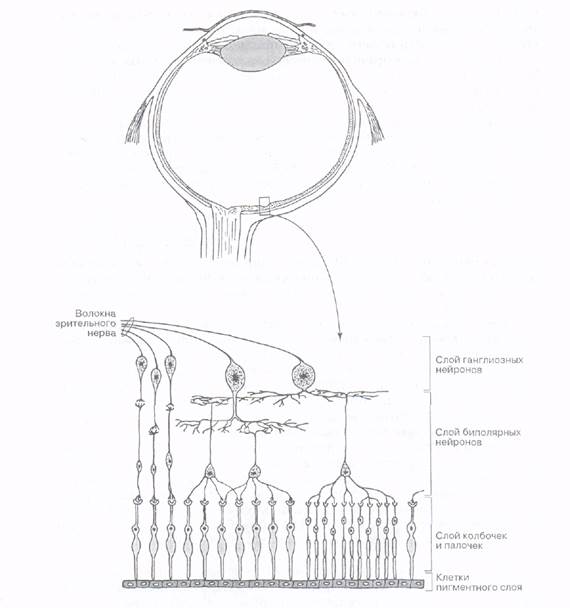
Рис. Нейронный состав сетчатки
Хрусталик (lens) - это прозрачная двояковыпуклая линза (то есть, по сути, увеличительное стекло) диаметром около 9 мм, имеющая переднюю и заднюю поверхности, которые переходят одна в другую в области экватора. Передняя поверхность хрусталика более плоская, чем задняя. Линия, соединяющая наиболее выпуклые точки обеих поверхностей (полюсы), называется осью хрусталика. Ее размеры колеблются в пределах 3,7-4,4 мм в зависимости от степени аккомодации. Коэффициент преломления хрусталика в поверхностных слоях равен 1,32, в центральных - 1,42. Хрусталик покрыт прозрачной капсулой, представляющей собой толстую базальную мембрану с множеством ретикулярных волокон. Под капсулой передняя половина хрусталика до его экватора покрыта однослойным плоским эпителием, задняя половина лишена эпителия. Эпителиальные клетки, расположенные вблизи экватора, являются ростковыми, они делятся, удлиняются, дифференцируются в хрусталиковые волокна и накладываются на периферические волокна позади экватора, в результате чего диаметр хрусталика увеличивается. В процессе дифференцировки ядро и органеллы исчезают, в клетке сохраняются лишь свободные рибосомы и микротрубочки. Ядро хрусталика образовано прозрачными волокнами призматической формы, состоящими большей частью из белка кристаллина. Эти волокна дифференцируются в эмбриональном периоде из эпителиальных клеток, покрывающих заднюю поверхность образующегося хрусталика, и сохраняются в течение всей жизни человека. Волокна склеены между собой веществом, чей индекс светопреломления аналогичен таковому волокон хрусталика.
Хрусталик как бы подвешен на ресничном пояске, то есть на цинновой связке. Между волокнами этого пояска расположены пространства пояска (петитов канал), сообщающиеся с камерами глаза. Волокна пояска прозрачны, они сливаются с веществом хрусталика и передают ему движения ресничной мышцы. При натяжении связки, то есть при расслаблении ресничной мышцы хрусталик уплощается (установка на дальнее видение), при расслаблении связки, то есть при сокращении ресничной мышцы, выпуклость хрусталика увеличивается (установка на ближнее видение). Это и называется аккомодацией глаза.
Стекловидное тело (corpus vitreum) заполняет пространство между сетчаткой сзади, хрусталиком и задней стороной ресничного пояска спереди. Оно представляет собой прозрачное аморфное межклеточное вещество желеобразной консистенции, его индекс светопреломления - 1,334. Стекловидное тело образовано тончайшими волокнами, составляющими его остов, между которыми находится жидкость. В состав стекловидного тела входит гигроскопический белок витреин и гиалуроновая кислота. На передней поверхности стекловидного тела имеется ямка, в которой располагается хрусталик.
Физические основы зрения человека. Зрение представляет собой восприятие электромагнитных излучений сравнительно узкого диапазона, то есть с длиной волны от 400 нм до 750 нм (при длине волны, равной 435 нм, возникает ощущение синего цвета, 546 нм - зеленого, 589 нм - желтого, 671 нм - красного). Для восприятия разной длины света в сетчатки имеются соответствующие зрительные пигменты, находящиеся в палочках или колбочках. Пигмент родопсин (преимущественно содержится в палочках) имеет максимум поглощения для излучения с длиной волны в 500 нм, йодопсин, хлоролаб и эритролаб (пигменты колбочек) - для излучений с длиной волной соответственно 560 нм, 546 нм и 671 нм. При этом орган зрения приспособлен к восприятию различного по интенсивности светового потока. Если яркость среды выразить в канделах (единица измерения интенсивности света) на м2, то ночью без луны и звезд яркость среды составляет 10-6 кд/м2, ночью со звездами -10-3 кд/м2, ночью при полной луне -10-1кд/м2, а в яркий солнечный день - 107 кд/м2. Таким образом, диапазон изменения интенсивности освещенности, воспринимаемой глазом, огромен (от 10-6 до 107 кд/м2).
Пигментный слой сетчатки. Он образован одним рядом эпителиальных клеток (пигментных эпителиоцитов), лежащих на базальной мембране. От внутренней поверхности пигментных клеток отходит по 8-10 цитоплазматических отростков («борода »), заполненных пигментом меланином, или фусцином (он синтезируется меланосомами эпителиоцитов), отделяющих друг от друга наружные сегменты палочек и колбочек. Благодаря меланину доходящий до сетчатки свет не рассеивается и не отражается, а полностью попадает на фоторецепторы, что способствует четкости зрительного восприятия. Эти же клетки поглощают избыточное количество квантов света при наличии чрезмерной освещенности сетчатки (при сильном освещении зерна пигмента перемещаются из эпителиальных клеток и заслоняют палочки и колбочки от яркого света). Кроме того, эпителиальные клетки пигментного слоя принимают участие в ресинтезе зрительного пигмента фоторецепторов после его обесцвечивания, а также (за счет фагоцитоза) в удалении отработанных наружных сегментов зрительных клеток. С помощью пигментных эпителиоцитов происходит перенос к фоторецепторам кислорода и питательных веществ. Нарушение всех этих функций пигментного слоя, которое наблюдается при отслойке сетчатки, вызывает дегенерацию фоторецепторов, что приводит к развитию слепоты.
У ночных животных между пигментными клетками и фоторецепторами расположен слой, отражающий свет. Он состоит из особых кристаллов или нитей. В результате отражения света от кристаллов у ночных животных при внешнем освещении светятся глаза. В этом случае на фоторецепторы действуют не только прямые лучи, но и отраженные, что в условиях слабой освещенности повышает возможность восприятия света.
Восприятие света осуществляется с участием фоторецепторов, или нейросенсорных клеток, которые относятся ко вторичночувствующим рецепторам. Это означает, что они представляют собой специализированные клетки, передающие информацию о квантах света на нейроны сетчатки, в том числе вначале на биполярные нейроны, затем на ганглиозные клетки, аксоны которых составляют волокна зрительного нерва; информация затем поступает на нейроны подкорковых (таламус и передние бугры четверохолмия) и корковых центров (первичное проекционное поле 17, вторичные проекционные поля 18 и 19) зрения. Кроме того, в процессах передачи и переработки информации в сетчатке участвуют также горизонтальные и амакриновые клетки. Все нейроны сетчатки образуют нервный аппарат глаза, который не только передает информацию в зрительные центры мозга, но и участвует в ее анализе и переработке. Поэтому сетчатку называют частью мозга, вынесенной на периферию.
Более 100 лет назад на основании морфологических признаков Макс Шультце разделил фоторецепторы на два типа - палочки (длинные тонкие клетки, имеющие цилиндрический наружный сегмент и равный ему по диаметру внутренний) и колбочки (обладающие более коротким и толстым внутренним сегментом). Он обратил внимание на то, что у ночных животных (летучая мышь, сова, крот, кошка, еж) в сетчатке преобладали палочки, а у дневных (голуби, куры, ящерицы) - колбочки. На основании этих данных Шультце предложил теорию двойственности зрения, согласно которой палочки обеспечивают скотопическое зрение, или зрение при низком уровне освещенности, а колбочки реализуют фотопическое зрение и работают при более ярком освещении. Следует, однако, отметить, что кошки прекрасно видят днем, а содержащиеся в неволе ежи легко приспосабливаются к дневному образу жизни; змеи, в сетчатке которых находятся главным образом колбочки, хорошо ориентируются в сумерках.
В настоящее время показано, что ночное, или палочковое, зрение имеет высокую чувствительность (в 1000 раз выше, чем колбочковое), низкую разрешающую способность (остроту) и является черно-белым (ахроматическим). Абсолютный порог зрения соответствует одновременному поглощению 6-7 фотонов сетчаткой, содержащей около 50 палочек. Палочковое зрение не цветное, так как все палочки содержат один зрительный пигмент - родопсин, и, следовательно, различия в длине волны воспринимаются как различия в интенсивности. Палочковое зрение неотчетливо потому, что конвергенция большого числа палочковых клеток на одно нервное волокно делает эффективную мозаику сетчатки довольно грубой. В целом, сегодня не вызывает сомнения, что палочковые фоторецепторы, расположенные в основном по периферии сетчатки (поэтому в сумерки лучше видны предметы, расположенные по сторонам) обеспечивают скотопическое и мезопическое зрение, то есть зрение при слабом дневном свете и при сумерках соответственно, и они предназначены для детектирования интенсивности света. Нарушение функции палочек вызывает расстройство сумеречного зрения - так называемую куриную слепоту: человек совершенно слепнет в сумерках, но днем зрение остается нормальным (обычно возникает при недостатке в пище витамина А).
Колбочковые фоторецепторы (три варианта) преимущественно сконцентрированы в области центральной ямки сетчатки (до 140 тыс. колбочек на 1 мм2). Они обеспечивают фотопическое зрение, т.е. зрение при обычном дневном свете, при котором человек способен различать цвета. Дневное, или колбочковое, зрение обладает более низкой абсолютной чувствительностью, чем палочковое в силу того, что колбочки по своей природе мене чувствительны и, кроме того, к одной ганглиозной клетке их сходится меньше, чем палочек. Однако уменьшение конвергенции приводит к повышению остроты зрения. Колбочковое зрение может быть цветным при наличии нескольких типов колбочек с различными зрительными пигментами, поглощающими в различных областях спектра. Поражение колбочек вызывает светобоязнь: человек видит при слабом свете, но слепнет при ярком освещении. В этом случае может развиться и полная цветовая слепота - ахромазия. Кроме того, отсутствие одного из видов колбочек приводит к нарушению цветоощущения, т.е. к дальтонизму.
Современные представления о процессах, происходящих в фоторецепторной клетке при воздействии света. В наружных сегментах палочек сетчатки человека содержится зрительный пигмент родопсин, или зрительный пурпур, максимум спектра поглощения которого находится в области 500 нм. Его молекулярная масса составляет 40 кД. Он относится к каротиноидам, т.е. к хромолипопротеинам. Поглощение света таким белком обеспечивается наличием хромофорной группы атомов, в качестве которой выступают альдегиды спиртов витамина А, их называют ретиналями. Во всех зрительных пигментах ретиналь всегда находится в 11-цисформе, у которой длинная часть цепочки изогнута и скручена в весьма нестабильную конформацию (такая форма называется 11- цис-ретиналем). В норме 11-цис-ретиналь связан с бесцветным белком опсином. Это соединение и является родопсином, который в той или иной модификации служит универсальным молекулярным медиатором фоторецепции у всех животных. Источником ретиналя в организме являются каротиноиды. Поэтому их недостаток приводит к дефициту витамина А, а как следствие, к недостаточному ре-синтезу родопсина, что в свою очередь является причиной нарушения сумеречного зрения («куриной слепоты»).
При поглощении кванта света родопсином происходит мгновенная цистрансизомеризация ретиналя, т.е. 11-цис-ретиналь выпрямляется и превращается в полностыо-транс-ретиналь. В результате этой реакции изомеризации, которая длится около 1 пс (10-12 с), происходит обесцвечивание родопсина, т.е. образование метародопсина II (последовательность событий: родопсин → прелюмиродопсин → люмиродопсин → метародопсин I → метародопсин II; при этом метародопсин I имеет темно-оранжевый цвет, а метародопсин II - светло-желтый.)
В свою очередь образовавшийся метародопсин II взаимодействует с трансдуцином (в соотношении 1 молекула метародопсина II на 500-1000 молекул трансдуцина). Трансдуцин представляют собой разновидность G-белка, т.е. гуанозинтрифосфат-связывающего белка, который широко представлен в различных клетках как посредник в передаче сигнала от мембранного рецептора к внутриклеточному эффектору (например, от в-адренорецепторов к кальциевому насосу). В комплексе с метародопсином II трансдуцин переходит в активное состояние и обменивает связанный с ним в темноте гуанозиндифосфат (ГДФ) на гуанозинтрифосфат (ГТФ). В свою очередь активированный трансдуцин, связанный с молекулой ГТФ, активирует фермент фосфодиэстеразу (ФДЭ) - в соотношении 1 молекула трансдуцина - 1 молекула ФДЭ. Активированная ФДЭ с высокой скоростью разрушает молекулы цГМФ, т.е. циклического гуанозинмонофосфата (1 молекула ФДЭ разрушает несколько тысяч молекул цГМФ), что приводит к существенному снижению концентрации свободного цГМФ в цитоплазме наружного сегмента рецептора. Так как именно цГМФ держит натриевые и кальциевые каналы наружного сегмента фоторецептора в открытом состоянии (это происходит в темноте), то снижение концентрации свободной цГМФ приводит к закрытию натриевых и кальциевых каналов, что в свою очередь порождает гиперполяризацию (т.е. генерацию рецепторного потенциала). Эта гиперполяризация тормозит выделение в пресинаптическом окончании фоторецепторной клетки глутамата, который в условиях темноты вызывал гиперполяризацию биполярной клетки. Тем самым происходит возбуждение биполярной клетки, которое, в свою очередь, передается на ганглиозную клетку (за счет выделения возбуждающего медиатора из пресинаптического окончания биполярной клетки, функцию которого, вероятнее всего, выполняет ацетилхолин). В ответ на генераторный потенциал ганглиозная клетка генерирует потенциалы действия, бегущие по зрительному нерву в подкорковые и корковые зрительные центры.
В связи с тем, что в наружном сегменте фоторецептора при действии света снижается содержание свободного кальция (из-за закрытия кальциевых каналов), это вызывает активацию фермента гуанилатциклазы, благодаря чему усиливается образование цГМФ из ГТФ. Повышение внутриклеточного содержания цГМФ вызывает открытие натриевых и кальциевых каналов в наружном сегменте фоторецептора, что приводит к деполяризации и восстановлению продукции глутамата. Одновременно накопление
цГМФ нарушает взаимодействие метародопсина II с трансдуцином. Таким образом, за короткий промежуток времени фоторецептор восстанавливает свою способность повторно отвечать на световой сигнал.
Отщепившийся от трансдуцина метародпосин II распадается на 11-транс-ретиналь и опсин. В последующем (в условиях темноты) происходит восстановление родопсина, т.е. соединение белка опсина с 11-цис-ретиналем. Ресинтез родопсина осуществляется с участием пигментного эпителия сетчатки при наличии ретинола, т.е. витамина А, а также фермента редуктазы. Редуктаза, с одной стороны, способствует превращению 11-транс-ретиналя в ретинол, т.е. в витамин А, а с другой - катализирует реакцию изомеризации, т.е. образование из витамина А его альдегида (11- цис-ретиналя), который в условиях темноты соединяется с опсином. Так завершается процесс ресинтеза родопсина.
В наружных сегментах трех типов колбочек (сине-, зелено- и красночувствительных) содержится три типа зрительных пигментов, максимумы спектров поглощения которых находятся в синей (435 нм), зеленой (546 нм) и красной (671 нм) частях спектра. Красный колбочковый пигмент получил название «йодопсин». Структура йодопсина близка к родопсину. Но в йодопсине 11-цис-ретиналь соединен с другим белком, который отличается от опсина палочек. Степень поглощения света родопсином и йодопсином различна. Родопсин максимально поглощает лучи в сине-зеленой части спектра. Эти лучи в темноте кажутся наиболее яркими. Йодопсин в наибольшей степени поглощает желтый свет.
Теории цветоощущения. Существует две основных теории цветового зрения - трехкомпонентная теория М.В. Ломоносова, Т. Юнга, Г. Гельмогольца и оппонентная теория цветоощущения Э. Геринга (или теория противоположных, или контрастных цветов). Вероятнее всего, что каждая из них объясняет последовательную работу двух механизмов цветовосприятия, а именно первичного (цветоприемного) механизма, связанного с функцией колбочек сетчатки, и вторичного (цветокодирующего) механизма, обусловленного обработкой зрительной информации в нейронах сетчатки, подкорковых и корковых центрах зрения.
Согласно трехкомпонентной теории, цветовое восприятие обеспечивается тремя типами колбочек с различной спектральной (цветовой) чувствительностью. Одни колбочки обладают максимумом возбуждения при их освещении красно-оранжевым (671-700 нм) светом, вторые - при освещении зеленым (546 нм) светом, а третьи - при освещении сине-фиолетовым (397-435 нм) светом. Всякий цвет оказывает действие на все три цветоприемных элемента, но в разной степени. Отметим, что до настоящего времени дискутируется вопрос о колбочках, чувствительных к зеленому цвету: по мнению ряда исследователей, они более чувствительны к желтому цвету, т.е. являются «желтоузнающими».
Разновидностью трехкомпонентной теории цветоощущения (менее популярной, чем предыдущая точка зрения) является представление о том, что все колбочки в сетчатке одинаковы и каждая из них содержит три различных светочувствительных вещества (цветоприемных элементов). Одно из них распадается при действии главным образом красного цвета, другое - зеленого, а третье - синего. Следовательно, в каждой колбочке имеется три приемника света и каждый из трех компонентов цветоощущения передается по своей системе сигналов (коду), отличной от других компонентов. Комбинацией излучений этих основных цветов можно получить все оттенки спектра, воспринимаемого зрением. Если одновременно и в одинаковой степени раздражаются все три типа цветоприемных элементов колбочек, то возникает ощущение белого цвета.
В целом, первый вариант трехкомпонентной теории цветоощущения подтвержден в опытах, в которых микроспектрофотометром измеряли поглощение излучений с разной длиной волны у одиночных колбочек сетчатки человека. Трехкомпонентная теория хорошо согласуется с представлением об аномалии цветовосприятия. Человек, имеющий все три вида колбочек, т.е. трихромат, обладает нормальным цветовосприятием. Отсутствие одного какого-либо типа колбочек (т.е. у дихроматов) приводит к нарушению цветоощущения, которое получило название частичной цветовой слепоты. Чаще это явление называют дальтонизмом - по имени известного деятеля Великой французский революции Д. Дальтона, носителя этого вида аномалии, который впервые описал ее. Дальтонизм встречается у 8% мужчин и намного реже у женщин.
Возникновение его связывают с отсутствием определенных генов в половой непарной у мужчин Х-хромосоме. Для диагностики дальтонизма, важной при профессиональном отборе, используют полихроматические таблицы, например, таблицы Е. Рабкина. Нарушение работы красного механизма (протанопия) встречается примерно у 1,1% мужчин, зеленого механизма (дейтеранопия) - у 1,5%, а синего механизма (тританопия) - у 0,001% мужчин. Протанопы («краснослепые») не воспринимают красный цвет, сине-голубые лучи кажутся им бесцветными; дейтеранопы («зеленослепые») не отличают зеленые цвета от темно-красных и голубых цветов, а тританопы не воспринимают лучи синего и фиолетового цвета. У 0,01% людей имеется лишь один вид колбочек (монохроматы). Крайне редко встречаются люди с полной цветовой слепотой (ахроматы). При ахромазии в результате поражения колбочкового аппарата сетчатки человек видит все предметы лишь в разных оттенках серого. Люди с частичной или полной цветовой слепотой не могут быть полноценными водителями транспорта, так как они не способны различать цвет огней светофоров и дорожных знаков.
Не исключено, что процессы фоторецепции в колбочках протекают по такому же типу, как в палочках. Функцию зрительного пигмента выполняют йодопсин (в наибольшей степени он поглощает свет с длиной волны 560 нм), а также хлоролаб (546 нм) и эритролаб (671 нм).
Представление о наличии вторичного - цветокодирующего механизма - первоначально было постулировано Э.Герингом как попытка объяснить ряд феноменов цветового зрения, в том числе феномен одновременного или последовательного контраста. В частности, Э.Геринг выдвинул представление о так называемых оппонентных цветах (красный и зеленый, синий и желтый, черный и белый), связывая это с синтезом или распадом зрительных пигментов.
В настоящее время установлено, что функцию различения цветов выполняют не только колбочки сетчатки, но и нейроны сетчатки (в том числе ганглиозные и горизонтальные клетки), нейроны наружного коленчатого тела и зрительной коры. Так, показано, что в сетчатке имеются нейроны, рецептивное поле которых устроено таким образом, что в его центре находятся колбочки, воспринимающие, к примеру, красный цвет, а на периферии - колбочки, которые нечувствительны к нему. Когда луч красного цвета возбуждает колбочки, находящиеся в центре рецептивного поля, нейрон возбуждается, в результате чего возникает ощущение красного. Если свет в основном поглощается колбочками на периферии рецептивного поля, то этот нейрон не возбуждается, что создает ощущение зеленого света. Это возможно при условии существования второго варианта рецептивного поля: в центре находятся колбочки, воспринимающие зеленый цвет, тогда возбуждение этих нейронов даст ощущение зеленого, если же кванты света воспринимаются в основном колбочками периферии, то нейрон не возбуждается, что вместе с другим типом объединения рецепторов дает ощущение красного. Синий и желтый цвет возникает в рецептивных полях, где в одном случае центр представлен колбочками, воспринимающими желтый цвет, а периферия - синий, и наоборот, рецептивные поля, где в центре концентрируются колбочки, воспринимающие синий цвет, а на периферии - желтый. Ощущение черного и белого цветов возникает в результате такой организации, когда центр воспринимает все цвета (т. е. колбочки чувствительны ко всем цветам - это дает ощущение
белого цвета), а периферия - не чувствительна к свету. Таким образом, любой цвет может быть представлен совокупностью нейронов, воспринимающих основные цвета. В результате многочисленного объединения нейронов на более высоком уровне (латеральное коленчатое тело, кора) возникает ощущение всех оттенков цвета. Оппонентность отмечена также для биполярных и горизонтальных клеток С-типа: цвет различного спектрального состава вызывает либо деполяризацию этих клеток, либо гиперполяризацию. Показано, что в наружном коленчатом теле спектрально оппонентные нейроны (цветочувствительные нейроны) представлены очень широко - на их долю приходится около 67% от общего числа нейронов. В зрительной коре также выявлены цветочувствительные нейроны. Их рецептивные поля устроены таким образом, что центр возбуждается на длинноволновый стимул и тормозится коротковолновым; напротив, периферия возбуждается коротковолновым и тормозится длинноволновым стимулом.
Таким образом, трехкомпонентная теория цветовосприятия (колбочки трех видов) достаточно хорошо согласуется с оппонентной теорией. В определенной степени эти две теории подтверждаются явлением одновременного и последовательного цветового контраста. Явление одновременного цветового контраста проявляется в том, что при длительном наблюдении серый круг приобретает зеленый цвет; если же серый круг имеет ободок зеленого цвета, то при длительном наблюдении ободок приобретает красную окраску. Явление последовательного цветового контраста (последовательных цветовых образов) проявляется в том, что если долго смотреть на окрашенный предмет (например, на красный квадрат), а затем перевести взор на белую бумагу, то тот же предмет виден окрашенным в дополнительный (зеленый) цвет. Причина этого явления в цветовой адаптации, т. е. снижении чувствительности к этому цвету. Поэтому из белого света как бы вычитается тот, который действовал на глаз до этого, и возникает ощущение дополнительного цвета.
Дата добавления: 2018-06-28; просмотров: 639;