Транспозиції мобільних елементів
Переміщення (транспозиції) мобільних (інтерсперсних) елементів є ще одним особливим типом рекомбінації ДНК, яка відбувається в усіх організмів. Мобільні елементи можна поділити на два основні класи: ДНК-транспозони, що переміщуються за принципом "cut and paste", і елементи, переміщення яких відбувається шляхом синтезу РНК із наступним синтезом ДНК на РНК-матриці. Основними типами мобільних елементів, які належать до другого класу, є: LTR-ретропозони з довгими кінцевими повторами (Long Terminal Repeats); елементи LINE (Long INterspersed Elements); елементи SINE (Short INterspersed Elements).
Переміщення мобільних елементів є досить рідкою подією – у бактерій відбувається одна транспозиція на 105 – 106 клітин. Проте активність мобільних елементів має важливе біологічне значення. Вбудовування мобільних елементів у кодуючі частини генів очевидно приводить до мутацій, у регуляторні – до зміни транскрипційної активності. У процесі свого переміщення мобільний елемент може захопити той чи інший регуляторний елемент послідовності та вбудувати його в інший промотор. Переміщення генетичного матеріалу разом із мобільними елементами розглядається сьогодні як один із найважливіших еволюційних факторів.
ДНК-транспозони
Мобільні елементи цього типу часто містять один або два гени, що кодують один або два білки під спільною назвою транспозаза. Транспозаза здійснює каталіз хімічних реакцій, що забезпечують транспозицію елемента – його вирізання з донорного сайта та вбудовування в сайт-мішень. Крім генів транспозази мобільний елемент може містити інші гени, або навпаки – гени транспозази можуть бути пошкодженими. В останньому випадку транспозиція даного елемента відбувається з використанням транспозази, закодованої іншим ДНК-транспозоном.
Кодуюча частина транспозона фланкована невеликими повторами, котрі розпізнає транспозаза, вирізаючи фрагмент ДНК з тупими кінцями (розрізи у двох полінуклеотидних ланцюгах розташовані точно один напроти одного). Сайт-мішень – невелика специфічна послідовність ДНК, яка теж упізнається транспозазою і теж розрізається – але так, що в місці розрізу залишаються одноланцюгові 5′-кінцеві вирости (рис. 15). Далі транспозаза каталізує ковалентне приєднання 3′-кінців транспозона до цих виростів, прогалини заповнюються шляхом репатаривного синтезу.
Процес транспозиції (який, по суті, є одним із варіантів сайт-специфічної рекомбінації) залишає дволанцюговий розріз у місці, де знаходився транспозон. У випадку незалежної від реплікації транспозиції (нереплікативна транспозиція), цей розріз піддається репарації за механізмом негомологічного з’єднання кінців, тобто транспозон просто «стрибає» в інше місце. Але досить часто транспозиція відбувається під час реплікації (реплікативна транспозиція) – тоді розріз репарується за рекомбінаційним механізмом (див рис. 1): сестринська молекула ДНК використовується як матриця, і ділянка, що містила транспозон, відновлюється – транспозон розмножується.
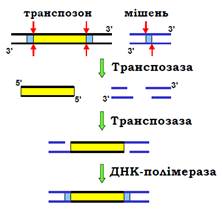
Рис. 15. Механізм переміщення ДНК-транспозона.
LTR-ретропозони
Кодуюча ділянка LTR-ретропозона (5 – 8 тис. пар основ) містить кілька генів: зворотної транскриптази (РНК-залежна-ДНК-полімераза), інтегрази (аналог транспозази), РНК-ази Н. На кінцях розташовані довгі кінцеві повтори (200 – 600 пар основ), які й надали назву цим мобільним елементам.
Процес переміщення копії LTR-ретропозона нагадує життєвий цикл ретровірусів. Першим етапом є транскрипція ретропозона: синтез мРНК РНК-полімеразою ІІ (рис. 16). Ця мРНК транспортується до цитоплазми, де піддається трансляції. Зворотна транскриптаза, якає продуктом цієї трансляції, здійснює синтез ДНК: мРНК використовується як матриця, 3′-кінець молекули тРНК – як праймер. Комплекс синтезованої ДНК-копії ретропозона з інтегразою повертається до ядра, де здійснюється інсерція цієї ДНК у геном.
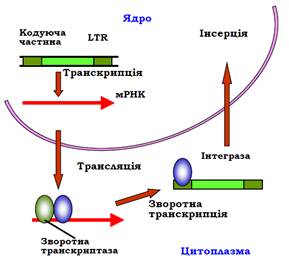
Рис. 16. Механізм переміщення LTR-ретропозона.
Дата добавления: 2018-03-01; просмотров: 769;