ГЛАВА 5. ГРУППА АБИОТИЧЕСКИХ ФАКТОРОВ
Общие сведения
Влияние климатических факторов (температура, влажность воздуха, осадки, ветер и др.) на организм всегда бывает совокупным. Однако изучение воздействия каждого отдельно взятого климатического фактора позволяет лучше понять его роль в жизни определенных видов или сельскохозяйственных культур и служит необходимой предпосылкой исследования воздействия всего комплекса климатических факторов. При оценке климатических факторов нельзя придавать исключительное значение лишь одному из них. Любой из названных компонентов климата в конкретных условиях может быть представлен по-разному: не только в количественном отношении, но и в качественном. Например, сумма годовых осадков для определенной местности может быть достаточно высокой, но распределение их в течение года неблагоприятно. Поэтому в отдельные периоды года (в вегетационные периоды) влага может выступать в роли минимум-фактора и тормозить рост и развитие растений.
Свет
Жизнь нашей планеты во всем ее разнообразии обусловлена энергией солнечной радиации. Эта же энергия приводит в движение сложный механизм воздушных и морских течений. Под ее воздействием происходит испарение воды и свершается непривычный влагооборот. Велика роль света в жизни растений. Только очень немногие виды растительных организмов осуществляют синтез, потребляя энергию не света, а химических связей неорганических соединений. Таковы серобактерии, железобактерии, нитрифицирующие бактерии. Свет необходим не только хлорофиллоносным растениям, но и гетеротрофам, например грибам-сапрофитам и даже паразитам.
У культур, особенно требовательных к свету, например риса, при недостаточной освещенности задерживается развитие. Формирование высокопроизводительных древостоев многих лесообразующих пород и плодовых насаждений также в значительной степени определяется интенсивностью солнечной энергии. Сахаристость свеклы прямо зависит от интенсивности лучистой энергии солнца в течение вегетационного периода. Известно, что у льна обыкновенного (Linum usitatissimum) и конопли посевной (Cannabis sativa) в условиях короткого светового дня синтезируется в тканях значительное количество масла, а в условиях длинного убыстряется формирование лубяных волокон. Реакция растений на длину дня и ночи проявляется в ускорении или задержке развития. Следовательно, действие света на растение избирательно и неоднозначно. Значение освещенности как экологического фактора для организма определяется продолжительностью, интенсивностью и длиной волн светового потока.
На границе земной атмосферы с космосом радиация составляет от 1,98 до 2 кал/см2 в 1 мин; Указанную величину называют солнечной постоянной. К поверхности Земли при разных погодных условиях доходит 42...70% солнечной постоянной. Солнечная радиация, проходя через атмосферу, претерпевает ряд изменений не только в количественном отношении, но и по составу. Коротковолновая радиация поглощается озоновым экраном, расположенным на высоте около 25 км, и кислородом воздуха. Инфракрасные лучи поглощаются в атмосфере водяными парами и диоксидом углерода. В результате нагревается воздух. Остальная часть лучистой энергии достигает поверхности Земли в виде прямой или рассеянной радиации (рис. 10). Совокупность прямой и рассеянной солнечной радиации составляет суммарную радиацию В ясные дни рассеянная радиация составляет от 1/3 до 1/8 суммарной радиации, тогда как в облачные дни рассеянная радиация составляет 100 %. В высоких широтах преобладает рассеянная радиация, под тропиками - прямая. Рассеянная радиация содержит в полдень желто-красных лучей до 60 %, прямая - 30...40 %.
Количество достигающей поверхности Земли радиации обусловлено географической широтой местности, продолжительностью дня, прозрачностью атмосферы и углом падения солнечных лучей. В ясные солнечные дни лучистая энергия, достигающая поверхности Земли, состоит на 45 % из видимого света (380...720 нм) и на 45 % из инфракрасного излучения, только 10 % приходится на ультрафиолетовое излучение. Значительное влияние на радиационный режим оказывает запыленность атмосферы. В некоторых городах вследствие ее загрязненности освещенность может составлять 15 % и менее освещенности за городом.
Освещенность на поверхности Земли варьирует в широких пределах. Все зависит от высоты стояния солнца над горизонтом, т. е. угла падения солнечных лучей, длины дня и условий погоды, прозрачности атмосферы. Интенсивность света также колеблется в зависимости от времени года и времени суток. Неравноценно в отдельных районах Земли и качество света, например соотношение длинноволновых (красных) и коротковолновых (синих и ультрафиолетовых) лучей. Как известно, коротковолновые лучи больше, чем длинноволновые, поглощаются и рассеиваются атмосферой. Поэтому в горных местностях всегда больше коротковолновой солнечной радиации.
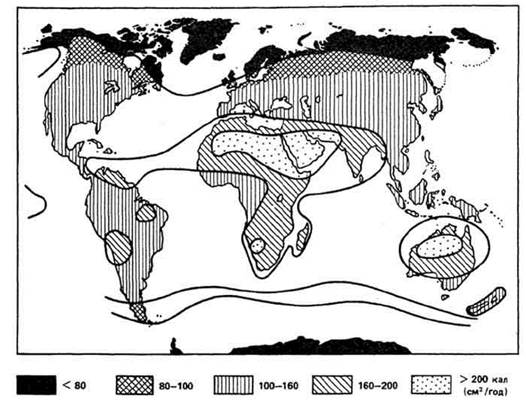
Рис. 10. Интенсивность солнечной радиации, падающей на поверхность Земли, по В. Лархеру
Поскольку фотосинтетически активная радиация (ФАР) представлена участком спектра между длиной волны 380 и 710 нм и максимальна в области оранжево-красных лучей (600...680 им), то естественно, что коэффициент использования растениями рассеянной радиации выше. Вследствие увеличения длины дня свет даже в высоких северных широтах не ограничивает жизнедеятельности растений. Л. Иванов рассчитал, что даже на Шпицбергене солнечной радиации достаточно (20 000 кДж на 1 га) для получения некоторою урожая сухой растительной массы.
У разных видов растений и растительных группировок потребность в свете неодинакова, иначе говоря, для нормальной вегетации им необходимо и неодинаковое световое довольствие (£,), т. е. доля в процентах полной ФАР. Это позволяет выделить в отношении потребности к свету три экологические группы растений:
· световые растения, или гелиофиты (от греч. helios - солнце + phyton), - Lопт = 100 %, £мин = 70 %, это растения открытых пространств, например ковыли (Stipa), большинство культивируемых растений (сахарная свекла, картофель и др.);
· теневыносливые растения, или гемисциофиты, могут расти при L = 100 %, но переносят и большое затенение; ежа сборная (Dactylis glomerata), например, способна вегетировать при диапазоне L от 100 до 2,5 %;
· теневые растения, или сциофиты (от греч. skia - тень), не переносят полного освещения, их Lмакс всегда меньше 100 %, это кислица обыкновенная (Oxalis acetosella), седь-мичник европейский (Trientalis europaea) и др.; ввиду особой структуры листьев сциофиты при слабой интенсивности света способны ассимилировать диоксид углерода не менее эффективно, чем листья гелиофитов при L = 100 %.
Московский растениевод А. Дояренко установил, что для большинства сельскохозяйственных травянистых растений коэффициент использования света для фотосинтеза составляет 2...2,5 %, но есть и исключения:
· свекла кормовая - 1,91
· вика - 1,98
· клевер - 2,18
· рожь - 2,42
· картофель - 2,48
· пшеница - 2,68
· овес - 2,74
· лен - 3,61
· люпин - 4,79
Из растительных сообществ лесные наиболее активно трансформируют состав солнечного света, и до поверхности почвы доходит очень малая часть первоначальной солнечной радиации. Известно, что листовая поверхность древостоя поглощает около 80 % падающей ФАР, еще 10 % отражается и только 10 % проникает под полог леса. Следовательно, полная и проникшая сквозь полог древесных растений радиация различается не только количественно, но и качественно.
Сциофиты и гелиоциофиты, обитающие под пологом других растений, довольствуются лишь долей полного освещения. Так, если у кислицы максимум интенсивности фотосинтеза достигается при 1/10 полного дневного освещения, то у светолюбивых видов он наступает примерно при 1/2 этого освещения. Световые растения менее приспособлены к существованию при слабом освещении, чем теневые и теневыносливые. Нижний предел, при котором могут расти лесные зеленые мхи, - 1/90 полного дневного освещения. В дождевых тропических лесах встречаются еще более сциофильные виды, вегетирующие при 1/120 полного освещения. Удивительны в этом отношении некоторые мхи: шистостега перистая (Schistostega pennaia) и др. - это растения темных пещер, вегетирующие при 1/2000 полного освещения.
Каждая географическая местность характеризуется определенным световым режимом. Важнейшими элементами светового режима, определяющими направление адаптации растений, являются интенсивность радиации, спектральный состав света, продолжительность освещения (длина дня и ночи). Продолжительность солнечного дня постоянна лишь на экваторе. Здесь день, как и ночь, длится 12 ч. Длительность солнечного дня в течение летнего периода возрастает от экватора в направлении обоих полюсов; на полюсе, как известно, целое лето длится полярный день, а зимой - полярная ночь. Реакцию растения на сезонные изменения длины дня и ночи называют фотопериодизмом.
Растениеводы давно обратили внимание на то, что сельскохозяйственные растения разного происхождения неодинаково реагируют на длину светового дня. В зависимости от этой реакции одни виды выделили как растения длинного дня, другие - короткого, третьи - как не реагирующие заметно на длину дня. Общеизвестно, что в условиях длинного дня формируется высокий урожай пшеницы, ржи, овса (Avena sativa) и ряда кормовых злаков; к растениям длинного дня относятся также картофель, цитрусовые и ряд других овощных и плодовых культур. Продолжительное освещение указанных растений вызывает более быстрое прохождение фаз развития плодов и семян. С другой стороны, у растений короткого дня, например проса (Panicum miliaceum), сорго (Sorghum сегпиит), риса, скорость прохождения этапов развития при длительном освещении замедляется. Сокращение периодов развития достигается укорачиванием времени освещения.
Указанные особенности необходимо учитывать при интродукции сельскохозяйственных растений. Виды низких широт (южные растения) часто бывают растениями короткого дня. При интродукции в высокие широты, т. е. в условия длинного дня, они развиваются медленно, часто не вызревают, а иногда даже не цветут, как, например, конопля. К данной группе можно отнести и топинамбур (Helianthus tuberosus). Таким образом, продолжительность дня и ночи может определять границы распространения и возможной интродукции отдельных видов: "южных" - на север, "северных" - на юг, К числу нейтральных в отношении длины дня относятся томат, виноград, гречиха посевная (Fagopyrum esculentum) и др.
В ходе изучения фотопериодизма и фотохимических реакций выяснено, что у растений длинного дня в весенне-летний период, когда в природе наблюдается длинный световой день, явно убыстряется рост. Однако во второй половине лета, когда солнечный день сокращается, ростовые процессы явно замедляются. В результате в условиях холодного климата у длиннодневных растений до наступления морозов не всегда успевает сформироваться комплекс покровных тканей - перидерма. Поэтому длиннодневные многолетние культуры, возделываемые в условиях высоких широт, могут утратить зимостойкость, что нужно иметь в виду при подборе ассортимента растений для возделывания в этих местностях. Предпочтительнее в условиях длинного дня вводить однолетние культуры, не требующие перезимовки. Продвижению к северу некоторых других культур, например клеверов, препятствуют не зимние морозы, а характер фотопериодических реакций. Именно их характером можно объяснить тот парадоксальный факт, что морозостойкость клеверов и люцерны более высока в средней полосе европейской части России, чем в северной.
Свет оказывает формирующее действие на растения, что проявляется в размерах, форме и структуре (макро- и микроскопической) световых и теневых листьев (рис. 11), а также в ростовых процессах. Зависимость структуры листа (побега) от света не всегда прямая; листья (побеги), развивающиеся весной, формируются в соответствии в освещением не текущего года, а прошлого, т. е. когда закладывались почки. И. Серебряков (1962) считал, что световая структура листа детерминирована уже в почке. Листья сохраняют данную структуру достаточно устойчиво даже при перенесении световых побегов в затенение. Большая высота, колоннообразная форма стволов, высокое расположение крон (очищение от сухих ветвей) характеризуют светолюбивые растения.
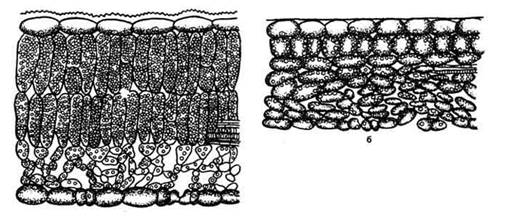
Рис. 11. Поперечные срезы листьев сирени (род Syringa): а - светового; б - теневого
Одна из реакций светолюбивых растений - затормаживание роста надземных побегов, что приводит в одних случаях к сильной ветвистости, в других - к розеточности. Отличает растения упомянутой группы и ряд других изменений структуры: мелколистность, увеличение мощности наружной стенки эпидермы и ее выростов (трихом и эмергенцев), кутикулярного слоя и т. д. (рис. 12).
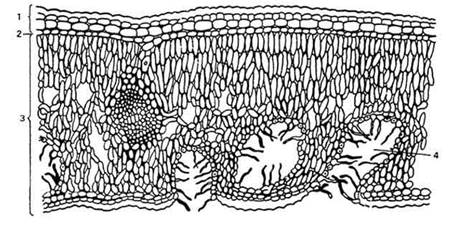
Рис. 12. Поперечный срез листа светолюбивого растения олеандра (Nerium oleander):
1 - двухслойная эпидерма с кутикулой; 2 - гиподерма; 3 - изопалисадный мезофилл; 4 - углубления на нижней стороне листа (крипты) с устьицами и волосками
Один из примеров приспособления растений к свету - ориентировка листовой пластинки по отношению к солнечным лучам. Различают три способа ориентировки:
· листовая пластинка ориентирована горизонтально, т. е. перпендикулярно солнечным лучам; в этом случае максимально улавливаются лучи, когда солнце находится в зените;
· листовая пластинка ориентирована параллельно солнечным лучам, т. е. расположена более или менее вертикально, в результате растение лучше усваивает солнечные лучи в утреннее и предвечернее время;
· листовые пластинки расположены по побегу диффузно, как у кукурузы, - то вертикально, то горизонтально, поэтому солнечная радиация улавливается достаточно полно в течение всего светового дня.
Имеющиеся научные данные позволяют считать, что растения высоких широт, где преобладает низкое солнцестояние, чаще имеют вертикальную ориентацию листьев. При организации смешанных посевов, например кормовых трав, нужно обязательно учитывать структуру побегов компонентов посевов. Удачное сочетание кормовых трав с разной ориентацией листьев обеспечит больший урожай фитомассы.
Как уже отмечалось, в зависимости от недостатка или избытка освещенности многие растения способны размещать листья в плоскостях, перпендикулярной и параллельной направлению солнечных лучей, образуя так называемую листовую мозаику. Листовая мозаика образуется в результате рационального размещения неодинаковых по величине не только листовых пластинок, но и черешков. Типичную листовую мозаику можно наблюдать в фитоценозах с участием клена остролистного, липы мелколистной (Tilia cordata), вяза гладкого (Ulmus laevis), ильма горного (Ulmus glabra) и других древесных пород. Четко прослеживается листовая мозаика у многих растений с горизонтальным размещением ветвей, например у плюща обыкновенного (Hedera helix) и многих травянистых растений (рис. 13).

Рис. 13. Листовая мозаика у плюща (Hedera helix)
Компасные растения явно избегают сильного освещения. Их листовая пластинка расположена не перпендикулярно солнечным лучам, как у розеточных растений, а параллельно, как у эвкалиптов или у латука дикого (Lactuca serrtola), что предохраняет листья от перегрева в условиях избыточной солнечной радиации. Тем самым обеспечивается и благоприятное прохождение фотосинтеза и транспирации.
Существует целый ряд других адаптивных приспособлений, как структурных, так и физиологических. Иногда подобные приспособления носят явно сезонный характер, что хорошо иллюстрирует, например, сныть обыкновенная (Aegopodium podagrata). В типичном местообитании (дубравы) на растении в течение вегетационного периода формируются два "поколения" листьев. Весной, когда почки деревьев еще не распустились и полог леса пропускает много света, образуется листовая розетка, ее листья по структуре (микро- и макроскопической) явно световые.
Позднее, когда развивается густой полог леса и до поверхности почвы доходит лишь 3…4 % лучистой энергии, появляется второе "поколение" листьев, явно теневых. Нередко у одного отдельно взятого растения можно наблюдать одновременно световые и теневые листья. Листья нижних ярусов кроны шелковицы черной (Morus nigra) крупные, лопастные, тогда как верхние ярусы кроны несут световые листья - более мелкие, лишенные лопастей. У лесообразующих пород подобным же образом формируется периферия кроны: в верхних ярусах - световые листья, внутри кроны - теневые.
Температура
Жизнедеятельность любого вида протекает в определенных интервалах температур. При этом прослеживаются зоны оптимума, минимума и максимума. В зоне минимума или максимума происходит затухание деятельности организма. В первом случае низкие температуры (холод), а во втором - высокие (жара) приводят к нарушению его жизненных процессов. За пределами крайних температур лежит летальная зона, в которой наступает необратимый процесс отмирания растения. Следовательно, температуры определяют границы жизни.
Вследствие неподвижного образа жизни высшие растения выработали большую выносливость к суточным и сезонным (годовым) колебаниям температур. Многие лесообразующие породы нашей тайги - сосна сибирская, лиственница даурская (Larix dahurica) и др. - выдерживают понижения температуры до - 50 °С и ниже и летнее тепло до 25 °С и выше. Годовая амплитуда достигает 75 °С, а иногда 85...90 °С. Виды растений, выдерживающие большие перепады температур, называют эвритермными (от греч. eurys + therme - тепло) в отличие от стенотермных.
Дифференциация тепла на нашей планете - основа широтной зональности и высотной поясности растительности и почв. Вследствие уменьшения от экватора к полюсам высоты солнцестояния и угла падения лучей изменяется количество тепла. Так, среднегодовая температура около экватора составляет 26,2 °С, возле 30 °с. ш. она уже равна 20,3 °С, а при 60° с. ш. снижается до - 1 °С.
Помимо среднегодовой температуры данной местности, важное значение в жизни организмов имеют наиболее высокая и наиболее низкая температуры (абсолютный максимум и абсолютный минимум), наблюдаемые в данной климатической зоне, а также средняя температура самого теплого и самого холодного месяца. Так, продолжительность вегетационного периода в тундре (т. е. выше 70° с. ш.) составляет всего полтора - два с половиной месяца при средней температуре 10...12 °С.
Тайга, иначе зона хвойных лесов, имеет вегетационный период три - пять месяцев, среднюю температуру 14.. Л6 °С. В южной части зоны, где преобладают хвойно-широколиственные леса, вегетация длится четыре-пять месяцев, средняя температура составляет 15... 16 °С. В зоне широколиственных лесов (40...50° с. ш.) вегетационный период - пять-шесть месяцев, средняя температура 16...18 °С. Резким контрастом описанным зонам выступает зона дождевых тропических лесов (0...15° с. и ю. ш.). Вегетационный период здесь круглогодичный со средней температурой 25...28 °С и часто не дифференцирован на сезоны. Исключительно важная особенность тропических районов в том, что разница между средними температурами самого теплого и самого холодного месяца менее контрастна, чем суточные колебания.
Рост растений непосредственно связан с температурным фактором. Зависимость отдельных видов от температуры колеблется в широких пределах. Четко различаются термофильные (от греч. therme + philia - любовь) растения и их антиподы - холодовыносливые, или криофильные (от греч. kryos - холод). А. Декандоль (1885) выделял группы гекистотермных, микротермных, мезотермных и мегатермных растений (от греч. gekisto - холод, mikros - малый, mesos - средний, megas - большой).
Перечисленные группы растений по отношению к температуре - комплексные, при их выделении учитывают и отношение растений к влаге. Дополнением к данной классификации можно считать выделение растений криофитов и психрофитов (от греч. psychros - холод + phyton) - гекистотермов и частично микротермов, требующих различных режимов увлажнения. Криофиты произрастают в холодных сухих условиях, а психрофиты - это холодостойкие растения влажных почв.
Не менее наглядно влияние температур на распространение отдельных видов растений и их группировок. Давно уже установлена связь географического распространения отдельных видов с изотермами. Как известно, виноград созревает в пределах изотермы со средней температурой в течение шести месяцев (апрель - сентябрь) 15 °С. Распространение дуба черешчатого на север ограничено годовой изотермой 3 °С; северная граница плодоношения финиковой пальмы совпадает с годовой изотермой 18... 19 °С.
В целом ряде случаев распространение растений обусловлено не только температурами. Так, изотерма 10 °С проходит с запада на восток через Ирландию, Германию (Карлсруэ), Австрию (Вена), Украину (Одесса). Названные местности имеют достаточно различный видовой состав природного растительного покрова и представляют возможность интродукции и возделывания разнообразного набора культур. В Ирландии часто не вызревают зерновые культуры. В Германии и Ирландии не вызревают многие тыквенные (арбузы - Citrullus vulgaris, дыни), хотя в открытом грунте произрастают камелии (Camella) и пальмы. В Карлсруэ в открытом грунте растут плющ и падуб (Ilex), иногда вызревает и виноград. В районе Одессы возделывают дыни и арбузы, но плющ и камелии не выдерживают низких температур зимы. Таких примеров можно привести много.
Таким образом, средние температуры в отрыве от других факторов среды не могут служить надежным показателем (индикатором) возможности интродукции и возделывания интересующей нас культуры. Суть в том, что разные виды растений характеризуются неодинаковой продолжительностью вегетационного периода. Поэтому в отношении температуры необходимо учитывать как продолжительность периода благоприятных температур для нормального развития растений, так и время наступления и продолжительность действия минимальных температур (то же в отношении максимальных).
В экологической и растениеводческой литературе для оценки тепловых ресурсов вегетационного периода широко используют сумму активных температур. Она служит хорошим показателем при оценке потребности растений в тепле и дает возможность определить район возделывания той или иной сельскохозяйственной культуры. Сумма активных температур состоит из суммы положительных среднесуточных температур за период, когда она выше 10 °С. В районах, где сумма активных температур равна 1000...1400 °С, можно возделывать ранние сорта картофеля, корнеплоды; где эта сумма достигает 1400...2200 °С, - хлебные злаки, картофель, лен и др.; сумма активных температур 2200...3500 °С соответствует зоне интенсивного плодоводства; при сумме этих температур более 4000 °С успешно возделывание субтропических многолетников.
Организмы, жизнедеятельность которых и температура тела зависят от тепла, поступающего из окружающей среды, называют пойкилотермными (от греч. poikilos - различный). К ним относят все растения, микроорганизмы, беспозвоночных животных и некоторые группы хордовых. Температура тела пойкилотермных организмов зависит от внешней среды. Вот почему экологическая роль тепла в жизни всех систематических групп растений и названных групп животных имеет первостепенное значение. Высокоорганизованных животных (птиц и млекопитающих) относят к группе гомойотермных (от греч. homoios - одинаковый), у которых температура тела постоянная, поскольку поддерживается за счет собственного тепла.
Известно, что протопласт клеток живых организмов способен нормально функционировать в интервале температур 0...50 °С. Только организмы, которые имеют специальные приспособления, могут выдерживать указанные экстремальные температуры в течение длительного времени. Физиологи установили оптимальные и критические температуры дыхания и других функций. Оказывается, нижний предел температуры дыхания у зимующих органов (почки, хвоя) - 20... - 25 °С. При повышении температуры интенсивность дыхания возрастает. Температуры свыше 50 °С разрушают белково-липидный комплекс поверхностного слоя цитоплазмы, что приводит к потере клетками осмотических свойств.
В некоторых районах России периодически наблюдается массовая гибель растений от слишком низких температур. Катастрофическое действие последних в наибольшей степени сказывается в малоснежные зимы преимущественно на озимых хлебах. Губительны и внезапные похолодания весной, когда растения трогаются в рост (поздневесенние заморозки). Нередко от холода гибнут не только интродуцированные вечнозеленые древесные, например цитрусовые, но и листопадные растения. Н. Максимов, изучая механизм действия низких температур, пришел к выводу, что причина гибели растений объясняется обезвоживанием цитоплазмы. В межклетниках ткани происходит кристаллизация воды. Кристаллы льда оттягивают воду из клеток и механически повреждают органеллы клеток. Критический момент наступает именно с появлением кристаллов льда внутри клеток.
Выделены природные группы морозоустойчивых растений. К ним можно отнести хвойные вечнозеленые деревья и кустарники, а также бруснику (Vaccinium vitis-idea), вереск и др. Среди травянистых многолетников также выявлено немало морозоустойчивых растений, способных переживать суровую зиму. В период зимнего покоя растения могут выдерживать очень низкие температуры. Так, побеги смородины черной (Ribes nigrum) при медленном снижении температуры до - 253 °С (температура, близкая к абсолютному нулю) могут сохранять жизнеспособность.
Большинство видов растений характеризуются индивидуальными реакциями на температуру. Так, весной прорастание зерновок ржи начинается при 1...2 °С, семян клевера лугового (Trifolium pratense) - при 1 °С, люпина желтого (Lupinus luteus) - при 4...5, риса - при 10...12 °С. Оптимальные температуры для вызревания семян этих культур составляют соответственно 25, 30, 28, 30...32 °С.
Для нормального роста и развития растений необходима соответствующая температура окружающей среды для надземных и подземных органов. Например, лен нормально развивается при температуре корня примерно в два раза ниже (10 °С), чем надземных органов (22 °С). В ходе онтогенеза потребность растений в тепле заметно меняется. Значительно варьирует температура органов тела растения и в зависимости от местонахождения (почва, воздух) и ориентировки по отношению к солнечным лучам (рис. 14). Экспериментально установлено, что прорастание семян рапса (Brassica napus), сурепицы (В. campestrts), пшеницы, овса, ячменя, клевера, люцерны и других растений наблюдается при температуре 0...2 °С, тоща как для появления всходов требуются более высокие температуры (3...5 °С).
Рис. 14. Температура (°С) разных органов растений: А - новосиверсии (Novosiversia glacialis), по Б. Тихомирову; Б - пролески сибирской (Scilla sibiriati, по Т. Горышиной, а - подстилка, б - почва
На многих видах континентальных растений благоприятно сказывается суточный термопериодизм, когда амплитуда ночных и дневных температур составляет 5... 15° С. Суть его заключается в том, что многие растения более успешно развиваются при пониженных ночных температурах. Например, томаты лучше развиваются, если дневная температура воздуха достигает 26° С, а ночная температура 17...18° С. Опытные данные свидетельствуют также, что растениям умеренных широт для нормального онтогенетического развития необходимы также осенние пониженные температуры - сезонный термопериодизм.
Температурный фактор влияет на растения на всех этапах их роста и развития. Причем в разные периоды каждый вид растений нуждается в определенных температурных условиях. Для большинства однолетних растений, например ячменя, овса и других, прослеживается общая закономерность: на ранних этапах развития температура должна быть ниже, чем на более поздних.
Мегатермные растения тропического происхождения, например сахарный тростник (Saccharum officinarum), нуждаются в высоких температурах в течение всей жизни. Наибольшей выносливостью к сверхвысоким температурам отличаются растения жарких и сухих районов - эуксерофиты, а также многие суккуленты, например Кактусовые и Толстянковые (Crassulaceae). Это свойственно и растениям засоленных, особенно сульфидами и хлоридами, почв. Указанные виды, как показал еще X. Люденгорд (1925, 1937), сохраняют жизнеспособность даже при 70 °С. Хорошо переносят высокие температуры сильно обезвоженные семена и плоды. Именно на данном свойстве основан известный метод борьбы с возбудителем пыльной головни пшеницы (Ustilago trtttci). При термообработке пораженных семян гриб, будучи стенотермным, погибает, тоща как зародыш зерновки остается жизнеспособным.
Труднее решить вопрос о влиянии температуры на изменение структуры самого растения, его морфологию. Наблюдения в природе и экспериментальные сведения дают различные объяснения. В самом деле, такое приспособление, как сильное опушение почечных чешуи и листьев, представляется комплексным, оно служит защитой не только от яркого света, но и от высоких температур, а также от излишнего испарения влаги. Яркий блеск глянцевитых листьев, параллельное расположение листовой пластинки к солнечным лучам, войлочное опушение - все это, несомненно, предотвращает перегрев листа, а также излишнюю транспирацию.
Основатель экологии растений Е. Варминг (1895) наглядно продемонстрировал влияние температуры на формирование приземистых и розеточных форм растений в Арктике и в высокогорьях альпийского и субнивального поясов, т. е. у самой границы вечных снегов. Речь идет не только о травянистых бесстебельных, розеточных вроде девясила корнеглавого (Inula rhizocephala), но и о древесных жизненных формах - березе карликовой, можжевельнике туркестанском (Juniperus turcestanica), кедровом стланике и пр. Стелющиеся и подушечные формы растений, например минуартия арктическая (Minuartia arctica), наиболее приспособлены к условиям жизни у самой поверхности почвы под прикрытием снежного покрова. Когда снега нет, в припочвенном слое воздуха на высоте до 15...20 см сохраняется наиболее высокая температура и сила ветра минимальна. К тому же внутри "подушки", формируемой растением, создается особый микроклимат, и колебания температур здесь гораздо менее выражены, чем вне ее. На развитие приземистых форм температурный фактор может действовать и непосредственно, и косвенно - вследствие нарушения водоснабжения и минерального питания.
Наиболее велика роль прямого влияния температур в процессе геофилизации растений. Под геофилизацией понимают погружение нижней (базальной) части растения в почву (сначала гипокотиля, затем эпикотиля, первого междоузлия и т. д.). Данный феномен свойствен преимущественно покрытосеменным растениям. Именно в ходе их исторического развития геофилизация играла видную роль в трансформации жизненных форм от деревьев до трав. С погружением в почву основания побегов интенсивно развивается система придаточных корней, корневищ, столонов и других органов вегетативного размножения. Геофилизация была необходимой предпосылкой появления разнообразных подземных органов растений, особенно органов вегетативного размножения. Указанное дало покрытосеменным большие преимущества в борьбе за существование, за господство на континентах Земли.
В онтогенезе многих покрытосеменных геофилизация растений осуществляется при помощи особых втягивающих (контрактильных) корней. Интересные экспериментальные исследования по геофилизации провел П. Лисицын. Он выяснил, что втягивание в почву базальной части растения распространено гораздо шире, чем ранее предполагалось (рис. 15). У озимых сельскохозяйственных культур геофилизация улучшает условия зимовки, у яровых, например гречихи, - условия водоснабжения.
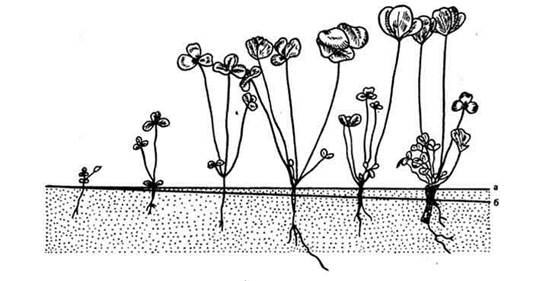
Рис. 15. Геофилизация (втягивание в почву) подсемядольного колена клевера лугового (Trifolium pratense), по П. Лисицину: а - поверхность почвы; б - глубина втягивания
Вода
Все процессы жизнедеятельности на уровнях клетки, ткани, организма немыслимы без достаточного водоснабжения. Органы растении обычно содержат 50...90 % воды, а иноща и больше. Вода - обязательный компонент живой клетки. Обезвоживание организма влечет замедление, а затем и прекращение жизненного процесса. Максимальное обезвоживание при сохранении жизни и обратимости нормальных жизненных процессов наблюдается в спорах и семенах. Здесь содержание воды падает соответственно до 10 и 12 %. Холодоустойчивость, равно как и жароустойчивость растений, зависит от количества находящейся в них воды. С водой связано и почвенное питание растений (поступление и транспортирование азотистых и других минеральных веществ), фотосинтез, ферментные процессы. Продукты метаболизма растворяются и транспортируются в теле растения также при помощи воды.
Вода - одно из необходимых условий формирования растительной массы. Установлено, что 99,5 % воды, транспортируемой от корневой системы к листьям, поддерживает тургор и только 0,5 % ее тратится на синтез органического вещества. Для получения 1 г сухой растительной массы требуется 250...400 г воды и более. Соотношение вышеуказанных значений составляет транспирационный коэффициент. У разных видов и даже сортов растений этот показатель существенно различается. Существует закономерность: величина транспирационного коэффициента прямо пропорциональна сухости климата. Поэтому один и тот же сорт может иметь неодинаковый транспирационный коэффициент при выращивании в разных эколого-географических условиях.
Оптимум водного режима наблюдается в случаях, когда испарение воды в атмосферу не превышает поступления ее в тело растения из почвы. В ходе онтогенеза наступает этап, когда снабжение водой определяет все последующее развитие растения и урожай. У многих культивируемых растений эти фазы развития хорошо изучены. Критический этап развития у злаков - формирование цветков и соцветий. При неблагоприятных условиях водоснабжения часть бугорков конуса нарастания дегенерирует. Поскольку данный процесс необратим, формируются укороченные, слабоветвистые соцветия, содержащие мало цветков, а следовательно, и зерновок.
На протяжении миллионов лет непрерывной эволюции организмы приспосабливались к различным условиям жизни. Растения аридных районов, где климат исключительно сухой, имеют ярко выраженные ксероморфные (от греч. xeros - сухой, morphe - форма) признаки. Они позволяют снижать потерю влаги, которая в основном происходит в результате транспирации через устьичный аппарат, а также через водяные устьица (явление гуттации - от лат. gutta - капля). Значительный расход влаги происходит и через клетки эпидермы (кутикулярное испарение). Гуттация хорошо выражена у проростков злаков, картофеля, гречихи, у многих комнатных растений, например у алоказии (Alocasia macrorhiza) и др. Наиболее распространена гуттация у растений влажных тропиков и субтропиков.
Растения засушливых условий имеют разнообразные приспособления, предупреждающие потерю воды. У многих злаков листья свернуты в трубку, так что устьица оказываются внутри. Листья ксероморфных растений часто имеют толстый восковой налет или волоски. Органы транспирации (устьичный аппарат) у таких растений погружены в мезофилл, часто наблюдается у них редукция листьев до чешуек или превращение в колючки и шипы. При сильной редукции листьев функцию фотосинтеза берет на себя стебель. Многие сельскохозяйственные культуры, как травянистые, так и древесные, реагируют на нехватку почвенной влаги и подземных вод быстрым развертыванием корневой системы.
Водный баланс растения определяется разностью между поглощением и расходованием воды организмом. На водный баланс влияет целая серия условий среды: влажность воздуха, сумма и распределение осадков, обилие и высота стояния подземных вод, направление и сила ветра.
Расход воды растениями в значительной мере определяется относительной влажностью воздуха. В более влажном климате при прочих равных условиях растения расходуют меньше влаги на образование сухого вещества. В умеренной зоне продуктивность транспирации составляет около 3 г сухих веществ при расходе 1 л воды, С возрастанием влажности воздуха в семенах, плодах и других органах растений содержится меньше белков, углеводов и минеральных элементов. Кроме того, уменьшается синтез хлорофилла в листьях и стеблях, однако одновременно усиливается рост и тормозятся процессы старения. При высоком насыщении воздуха водяными парами хлеба созревают очень медленно, а иногда не дозревают вовсе. Влажность воздуха оказывает большое влияние на количество и качество урожая, работу сельскохозяйственных машин. При высокой влажности воздуха возрастают потери урожая при обмолоте и уборке, а также замедляются процессы послеуборочного дозревания семян, что снижает в конечном счете их сохранность.
В зависимости от отношения к влаге растения разделяют на две экологические группы: пойкилогидридные и гомойгидридные. Первые не имеют специальных механизмов для регулирования гидратуры (обводненности) своего тела; по характеру потери влаги они практически не отличаются от мокрой хлопчатобумажной ткани. К пойкилогидридным относят низшие растения, мхи, многие папоротники. Абсолютное большинство семенных растений гомойгидридны и имеют специальные механизмы (устьичные аппараты, трихомы на листьях и т. д.) для регулирования внутреннего водного режима. Пойкилогидридность среди покрытосеменных крайне редка и имеет, скорее всего, вторичное происхождение, т. е. является своеобразной адаптацией к ксерическому режиму. Редким примером пойкилогидридного покрытосеменного растения служит пустынная осока вздутая, или илак (Carex physoides).
Гомойгидридные растения по характерному для них водному режиму подразделяют на гидрофиты, гелофиты, гигрофиты, мезофиты, ксерофиты, ультраксерофиты.
Гидрофиты (от греч. hydor - вода + phyton) - водные растения, свободно плавающие или укореняющиеся на дне водоема или полностью погруженные в воду (иногда с плавающими на поверхности листьями или выставленными над водой соцветиями). Поглощение воды и минеральных солей осуществляется всей поверхностью растения. У плавающих гидрофитов корневая система сильно редуцирована и иногда теряет свои функции (например, у рясок). Мезофилл подводных листьев не дифференцирован, отсутствуют кутикула и устьица". Примерами гидрофитов служат валлиснерия (Vallisneria spiralis), элодея канадская (Elodea canadensis), рдест плавающий (Potamogeton natans), альдрованда пузырчатая (Aldrovanda vesiculosa), кувшинка белая (Nymphaea alba), кубышка желтая (Nuphar luteum) и др. Для перечисленных видов характерны сильное развитие воздухоносной ткани - аэренхимы, большое количество устьиц у плавающих листьев, слабое развитие механических тканей, иногда разнолистность.
Гелофиты (от греч. helos - болото) - водно-наземные растения, растущие как в воде на мелководьях, так и по переувлажненным берегам рек, водоемов; могут обитать и на обильно увлажненной почве в удалении от водоемов. Встречаются только в условиях постоянного и обильного водоснабжения. К гелофитам относят тростник обыкновенный; частуху подорожниковую (Alisma plantago-aquaucd), стрелолист стрелолистный (Saggitaria sagittifolia), сусак зонтичный (Butomus umbellatus) и др. Гелофиты могут выдерживать недостаток кислорода в почве.
Гигрофиты (от греч. hygros - влажный) - наземные растения, произрастающие в условиях повышенной влажности почвы и воздуха. Они характеризуются насыщенностью тканей водой до 80 % и выше, наличием водяных устьиц. Различают две экологические группы гигрофитов:
· теневые, произрастающие под пологом сырых лесов в разных климатических зонах, для них характерны водяные устьица - гидатоды, позволяющие поглощать воду из почвы и транспортировать минеральные элементы, даже если воздух насыщен водяными парами; к теневым гигрофитам относят недотрогу обыкновенную (Impattens noli-tangere), цирцею парижскую (Circaea lutetiana), кислицу обыкновенную;
· световые, произрастающие на открытых местообитаниях, вде почва и воздух постоянно влажные; к ним относят папирус (Cyperus papyrus), росянку круглолистную (Drosera rotundifolia), подмаренник болотный (Galium palustre), рис посевной, калужницу болотную (Caltha palustrts).
Для гигрофитов характерна слабая приспособленность к регуляции обводненности тканей, поэтому сорванные растения данной группы очень быстро вянут. Таким образом, гигрофиты из наземных гомойгидридных растений наиболее близки к пойкилогидридным формам. Гидрофиты, гелофиты и гигрофиты имеют положительный водный баланс.
Мезофиты (от греч. mesos - средний) - растения, приспособленные к жизни в условиях среднего водоснабжения. Они проявляют высокую жизнеспособность в условиях умеренно теплого режима и средней обеспеченности минеральным питанием. Могут переносить непродолжительную не очень сильную засуху. К указанной группе принадлежит подавляющее большинство возделываемых культур, а также растения лесов и лугов. Вместе с тем мезофиты настолько разнообразны по морфофизиологической организации и приспособленности к различным местообитаниям, что им трудно дать общее определение. Они составляют разнообразную гамму промежуточных растений между гигрофитами и ксерофитами. В зависимости от распространения в различных климатических зонах А. Шенников (1950) выделил следующие пять групп мезофитов: вечнозеленые мезрфиты влажных тропических лесов - деревья и кустарники[*], вегетирующие круглый год без резко выраженного сезонного перерыва; для них характерны крупные листья с гидатодами, часто такие листья имеют на конце острие, отводящее воду; кожистость, пониклость и расчлененность листьев обеспечивают их сохранность во время дождей (филодендрон - Philodendron, фикус - Ficus elastica и др.); верхние широкие и плотные листья растений группы адаптированы к яркому освещению, им свойственна толстая кутикула, хорошо выраженная столбчатая паренхима, достаточно развитые проводящая система и механические ткани;
зимне-зеленые деревянистые мезофиты, или тропофиты (от греч. tropos - поворот), - также преимущественно виды тропической и субтропической зон, но распространенные не в дождевых лесах, а в саваннах; сбрасывают листву и впадают в состояние покоя во время сухого летнего периода; имеют хорошо выраженные покровные комплексы - перидерму и корку; типичный представитель - баобаб;
летне-зеленые деревянистые мезофиты - растения умеренного климата, деревья и кустарники, сбрасывающие листья и впадающие в состояние покоя в холодный период года; к ним относят большинство листопадных деревьев холодной и умеренной зон; опадение листвы зимой служит приспособлением к уменьшению испарения в холодные месяцы, когда всасывание воды из почвы затруднено; большое значение для данной подгруппы мезофитов имеют покровные комплексы (перидерма и корка), а также приспособления для защиты почек от потери воды; все же зимой растения теряют значительное количество влаги; испарение идет главным образом через слабозащищенные листовые рубцы и почки;
летне-зеленые травянистые многолетние мезофиты - растения умеренного климата, надземные части которых обычно отмирают на зиму, за исключением защищенных почек возобновления; очень обширная группа; наиболее типичные представители - многолетние луговые травы (тимофеевка луговая - Phleum pratense, клевер луговой и др.) и лесные травы (ясменник душистый - Asperula odorata, копытень европейский и др.); для листьев характерен дифференцированный мезофилл, хотя у лесных растений (сциофитов и гемисциофитов) палисадная ткань часто не выражена; проводящие элементы развиты умеренно; эпидерма тонкая, кутикула имеется не всегда; механические ткани развиты в средней степени или слабо;
эфемеры и эфемероиды (от греч. ephemeros - однодневный) - однолетние (эфемеры) и дву- или многолетние (эфемероиды) растения, которые в засушливых условиях вегетируют короткий влажный период, в сухой сезон переходят в состояние покоя; например, растения пустынь и сухих степей: эфемеры - веснянка весенняя, бурачок маленький (Alissum minutum) и др.; эфемероиды - мятлик живородящий, или курчавый (Роа bulbosa subsp. vMparum) различные виды тюльпанов (Tulipa), гусиных луков (Gagea), ирисов (Iris), ферул (Ferula) и др.; характерно отсутствие структурных адаптации к недостатку влаги, но семена способны переносить сильное высыхание и высокие температуры; луковичным и клубнелуковичным эфемероидам свойственны контрактильные (втягивающие) корни, обеспечивающие втягивание почки возобновления под почву на неблагоприятный период.
Необходимо отметить, что не все ученые согласны с отнесением пустынных эфемеров и эфемероидов к группе мезофитов и причисляют их к ксерофитам (понимая последний термин очень широко).
Ксерофиты (от греч. xeros) - растения, приспособленные к жизни в условиях низкого водоснабжения. Переносят почвенную и атмосферную засуху, так как имеют разнообразные приспособления для жизни в условиях жаркого климата при очень малом количестве осадков. Важнейшая особенность ксерофитов - формирование морфофизиологических адаптации к губительному действию атмосферной и почвенной засухи. В большинстве случаев ксерофиты имеют приспособления, ограничивающие транспирацию: безлистность, мелколистность, летний листопад, опушение. Многие из них способны длительное время выдерживать довольно сильное обезвоживание, сохраняя жизнеспособность. На рисунке 12 был показан лист с приспособлениями, ограничивающими испарение.
В зависимости от структурных особенностей органов и тканей, способов регулирования водного режима различают следующие три типа ксерофитов.
Первый тип - эуксерофиты (от греч. еu - настоящий) , или склерофиты (от греч. skleros - твердый), или собственно ксерофиты; по внешнему виду это суховатые, жесткие растения. Даже в период полного обеспечения водой обводненность их тканей мала. Склерофиты отличаются высокой устойчивостью к завяданию - могут без заметного вреда для себя терять до 25 % влаги. Их цитоплазма остается живой при столь сильном обезвоживании, которое было бы губительно для других растений. Еще одна особенность эуксерофитов - повышенное осмотическое давление клеточного сока, позволяющее значительно увеличивать сосущую силу корней.
Прежде считали, что интенсивность транспирации склерофитов, как и других ксерофитов, очень мала, однако работы Н. Максимова (1926, 1944) показали, что при благоприятных условиях водоснабжения эти растения транспирируют более интенсивно, чем мезофиты, особенно в пересчете на единицу поверхностного листа. И. Культиасов (1982) подчеркивал, что, по-видимому, основная особенность ксерофитов в их высокой засухоустойчивости, зависящей от свойств цитоплазмы, а также в способности эффективно использовать влагу после дождя. Характерная "склерофитная" морфология (мощное развитие механических и покровных тканей, мелколистность и др.) имеет защитное значение при затруднениях в водоснабжении.
Корневая система у эуксерофитов очень разветвленная, но неглубокая (менее 1 м). К рассматриваемой группе относят много растений наших степей, полупустынь и пустынь: полыни (белоземельная Artemisia terrae-albae, Лерха - А lerchlana и др.), веронику седую (Veronica incana) и др.
Д. Колпинов (1957) выделил из эуксерофитов особую группу - стипаксерофиты (от лат. stipa - ковыль). В нее входят узколистные злаки типа ковылей, типчака (Festuca valesiaca). Растения группы отличаются мощной корневой системой, использующей влагу кратковременных ливней. Стипаксерофиты чувствительны к обезвоживанию и переносят только кратковременный недостаток влаги.
Второй тип ксерофитов - гемиксерофиты (от греч. hemi - наполовину) имеют глубокую корневую систему, достигающую уровня грунтовых вод (до 10 м и более), т. е. являются фреатофитами (см. ниже).
Третий тип ксерофитов - суккуленты (от лат. succulentus - сочный) в отличие от ксерофитов описанных выше типов имеют хорошо развитую водозапасающую паренхимную ткань. В зависимости от ее дислокации различают листовые и стеблевые суккуленты. Примерами первых служат агавы (Agava), алоэ (Aloe), очитки (Sedum) и др. У стеблевых суккулентов листья обычно редуцированы, и воду эти виды запасают в стеблях (кактусы и кактусовидные молочаи).
Корневая система суккулентов обычно поверхностная. Они отличаются способностью запасать воду, когда она в окружающей среде находится в избытке, длительно удерживать и экономно расходовать ее. Транспирация у суккулентов чрезвычайно мала. Для ее сокращения у растений существует ряд приспособительных черт в строении, в том числе своеобразие форм надземных частей, демонстрирующих "знание" законов геометрии. Известно, что у сферических тел (особенно у шара) наименьшее отношение поверхности к объему. Утолщение листьев и стеблей, т. е. приближение их к шаровидной или цилиндрической форме, - способ сокращения транспирирующей поверхности при сохранении необходимой массы. У многих суккулентов эпидерма защищена кутикулой, восковым налетом, опушением. Устьица немногочисленны и днем обычно закрыты. Последнее обстоятельство создает затруднения для фотосинтеза, поскольку поглощение диоксида углерода этими растениями может идти в основном ночью: доступ СO2 и света не совпадает во времени. Поэтому у суккулентов выработался особый путь фотосинтеза - так называемый "САМ-путъ", при котором источником СО2 частично служат продукты дыхания.
Реакция корневой системы на водоснабжение хорошо изучена у культивируемых растений. На рисунке 16 показана глубина проникновения в почву корневой системы озимой пшеницы при различном количестве осадков.

Рис. 16. Корневая система озимой пшеницы (род Triticum):
1 - при большом количестве осадков; 2 - при среднем; 3 - при малом
Существует особая классификация экологических групп растений с учетом использования ими грунтовой влаги, т. е. по источникам поглощения влаги из субстрата. В ней выделяют фреатофиты (от греч. phreatos - колодец) - растения, корневая система которых постоянно связана с водоносными горизонтами почв и материнских почвообразующих пород, омброфиты (от греч. ombros - дождь) - растения, питающиеся влагой атмосферных осадков, и трихогидрофиты (от греч. trichos - волос) - растения, связанные с капиллярной каймой грунтовых вод, находящихся в состоянии постоянной подвижности. Среди фреатофитов выделяют облигатные и факультативные; последние довольно близки к трихогидрофитам. Для фреатофитов характерно развитие глубоко проникающих подземных органов; у верблюжьей колючки (Alchagi) - до 15 м, у древовидных форм черного саксаула (Haloxylon aphyllum) - до 25, у среднеазиатских тамариксов (Tamarix) - 7, у тамариксов Северной Африки - до 30, у люцерны посевной (Medicago sativa) - до 15 м. Омброфиты имеют неглубоко залегающую, но сильно разветвленную систему подземных органов, способную поглощать атмосферную влагу в большом объеме почвы. Типичные представители группы - эфемеры и эфемероиды пустынь. Для трихогидрофитов характерна корневая система универсального типа, в ней сочетаются черты фреатофитов и омброфитов. Фреатофиты и трихогигрофиты часто относят к группе гемиксерофитов.
Водоснабжение растений осуществляется из двух источников: осадков и грунтовых вод. Из атмосферных осадков важнейшую роль играют дождь и снег. Град, роса, туман, иней, ожеледь занимают более скромную долю в водном балансе растений. Атмосферные осадки для растений - не только источник водоснабжения. Твердые атмосферные осадки, образуя снежный покров, предохраняют почву, а следовательно, надземные и подземные органы растений от низких температур. Снежный покров в экологическом плане существенно влияет на среду обитания растений и животных - создает запас почвенной влаги, существенно понижает испарение влаги растениями. Важное значение для сельскохозяйственных растений, а также для продуктивности пастбищ и сенокосов имеют распределение осадков по сезонам, их форма, сумма и интенсивность выпадения.
Дожди, которые дают большое количество осадков в короткое время (более 1…2 мм/мин), называют ливневыми, или ливнями. Ливни обычно сопровождаются сильными ветрами и оказывают негативное влияние на сельскохозяйственные угодья. Самое большое количество осадков на Кавказе и в s Восточной Европе вообще (до 2500 мм в год) и ливневых дождей в частности приходится на Черноморское побережье Кавказа - Аджарию и Абхазию. Однако сильные ливни (свыше 5 мм/мин) зарегистрированы и на Украине. В целом же с продвижением на север внутри континента количество осадков сначала повышается, достигая максимума в умеренной зоне, а затем снижается (не распространяется на приморские районы); есть закономерность в изменении и других климатических показателей (рис. 17).
Большие различия (рис. 18) по количеству осадков между отдельными регионами Земли наряду с температурным режимом создают пестроту экологических условий на планете. Самые влажные районы располагаются в верхнем течении р. Амазонки, на о-вах Малайского архипелага.
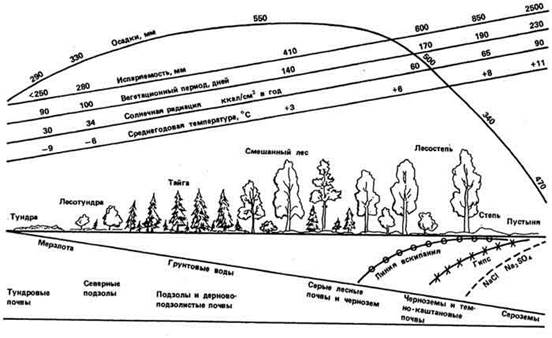
Рис. 17. Схематичный профиль европейской части России с севера на юг, по Г. Высоцкому
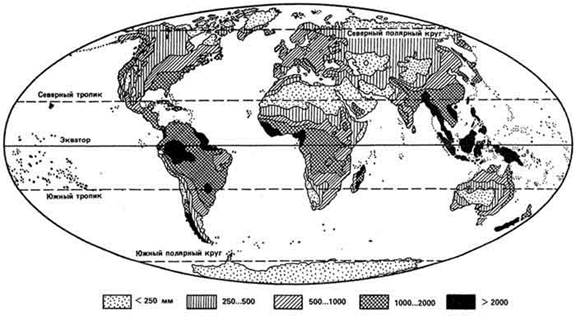
Рис. 18. Годовое распределение осадков по континентам
В зоне умеренного климата в местах, где наблюдаются частые оттепели, прослеживается гибель озимых от ледяной корки. После оттепелей талая снеговая вода, скопившаяся на полях в микропонижениях, замерзает и покрывает озимые культуры ледяной коркой. При этом происходит механическое давление льда, особенно губительно действующее на зоны кущения, одновременно ощущается недостаток кислорода.
В условиях чрезмерно толстого снежного покрова озимые могут выпревать. Причины выпревания заключаются в том, что снег обладает хорошими теплоизоляционными свойствами, поэтому в годы с мягкими зимами и с глубоким снежным покровом озимые преждевременно выходят из состояния покоя. В результате у растений заметно повышается интенсивность дыхания, однако, поскольку фотосинтез в данных условиях исключен, происходит резкое уменьшение запаса Сахаров. В результате растения истощаются, резко падает их резистентность. На ослабленных растениях бурно развивается паразитический гриб - снежная плесень (Fusartum nivale). Благоприятны для развития гриба долгая, холодная и влажная весна, глубокий снег и незамерзшая почва. Быстрое таяние снега и солнечная погода для снежной плесени губительны.
Толщина и плотность снежного покрова имеют важное значение для сельского, лесного, водного хозяйства. Рыхлый снег лучше защищает от охлаждения зимующие в почве растения. Плотность снега наименьшая при образовании снежного покрова, затем она постоянно возрастает и наибольшей становится в период таяния снега. Поэтому к весне защитное действие снежного покрова снижается. Части растений, не укрытые снегом, особенно в холодную и ветреную зиму, быстро теряют влагу и погибают. При температуре воздуха - 21 °С под снегом на поверхности почвы она составляет всего - 5 °С. Если снег выпадает рано и достаточно толстым слоем покрывает почву, она не промерзает, растения нормально растут и развиваются. Бывают зимы, когда под снежным покровом можно найти цветущие шафраны (род Crocus), любку двулистную (Platanthera bifolia) и другие растения.
В условиях суровой зимы высоких северных широт, а также в горах вырабатываются особые шпалерная и стланиковая формы одревесневающих растений. Даже крупноствольные деревья лесной зоны - ель сибирская, лиственница сибирская и другие - в условиях арктического климата трансформируются в стелющиеся формы.
Атмосферный воздух
Экологическое значение атмосферных осадков в жизни растений проявляется также в участии их как растворителя в подкормке минеральными веществами нижних ярусов древесных и травянистых растений. Во время дождя падающие капли насыщаются в воздухе летучими и парообразными веществами, последние вместе с каплей попадают на органы растений и поверхность почвы. Наряду с веществами, вымытыми из крон деревьев, и поглощаемыми летучими соединениями, выделяемыми растениями, в атмосферных осадках растворяются и смешиваются летучие и парообразные вещества, которые образуются в результате антропогенной деятельности, а также продукты жизнедеятельности почвенной микрофлоры.
Травянистые растения для данных экосистем нехарактерны, a эпифиты тропического леса относятся к подгруппам ксеромезофитов или гигромезофитов. Особенности их дислокации в кронах деревьев определяются микроклиматическими условиями.
Покрывающий Землю мощный слой воздуха (атмосфера) защищает живые организмы от мощного ультрафиолетового излучения и космической радиации, предотвращает резкие колебания температуры. Экологически не менее важны газовый состав атмосферы и перемещение воздушных масс (ветер и конвекционные потоки).
При характеристике газового состава воздуха обычно подчеркивают его постоянство. Почти во всех регионах земного шара сухой воздух тропосферы (нижнего слоя атмосферы) содержит около 78,1 % азота, 21 % кислорода, 0,032 % диоксида углерода, следы водорода, незначительное количество инертных газов. Наряду с постоянными компонентами в воздухе присутствуют газообразные составляющие, содержание которых варьирует в зависимости от времени и места: различные промышленные газы, аммиак, газообразные выделения растений и т. д.
Прямое экологическое влияние преобладающего в воздухе атмосферы свободного азота невелико; в данной форме указанный химический элемент оправдывает свое название, которое в переводе с греческого означает "не поддерживающий жизнь". Связанный азот служит важнейшим и обязательным компонентом всех биологических систем. Свободный атмосферный кислород не только поддерживает жизнь (дыхание), но и сам имеет биологическое происхождение (фотосинтез). Таким образом, ухудшение состояния зеленого мира нашей планеты может существенно сказаться на запасах свободного кислорода атмосферы.
Около 21 % выделяемого при фотосинтезе и содержащегося в воздухе кислорода потребляется растениями, животными и человеком в процессе дыхания. Взрослое дерево за сутки выделяет до 180 л кислорода. Человек потребляет в день при отсутствии физических нагрузок около 360 л кислорода, а при интенсивной работе - до 900 л. Легковой автомобиль на 1000 км расходует годичную норму кислорода, потребляемого человеком, а реактивный лайнер на перелет из Европы в Америку расходует 35 т кислорода.
Еще более зависит от жизнедеятельности различных организмов содержание в воздухе диоксида углерода. Важнейшими естественными источниками СO2 служат дыхание, брожение и гниение - на общую долю перечисленных процессов приходится 5.6,1 % поступления СO2 в атмосферу. Около 38 % диоксида углерода поступает в воздух из почвы ("почвенное дыхание"); 0,1 % - при извержении вулканов. Довольно существенным источником СO2 становятся лесные и степные пожары, а также сжигание топлива - до 0,4 %. Последний показатель постоянно растет: в 1970 г. вследствие антропогенной деятельности в воздух попало 0,032 % годового поступления СO2, по прогнозам ученых, к двухтысячному году доля рассматриваемого источника возрастет до 0,038...0,04 %.
Существенно сказывается деятельность человека и на темпах фиксации диоксида углерода в биосфере. Главным образом это объясняется чрезмерной вырубкой лесов и загрязнением Мирового океана. Растения при фотосинтезе связывают ежегодно 6...7 % СO2 воздуха, причем наиболее интенсивен процесс в лесных экосистемах. Дождевой тропический лес за год фиксирует 1...2 кг диоксида углерода на 1 м2, в тундрах и пустынях фиксируется лишь 1 % этого количества. Всего экосистемы суши фиксируют за год 20...30 млрд т СО2 Примерно столько же фиксируется фитопланктоном Мирового океана.
Возрастание содержания диоксида углерода в атмосфере имеет отрицательные экологические последствия в планетарном масштабе и проявляется в виде "парникового эффекта". В общих чертах указанный эффект можно охарактеризовать как постоянное потепление климата, вызываемое тем, что, подобно пленке в парнике, накопившийся в чрезмерном количестве СO2 препятствует оттоку длинноволнового теплового излучения от поверхности Земли, свободно пропуская при этом солнечные лучи. Конкретные проявления "парникового эффекта" неодинаковы в различных регионах. В одном случае это небывалые засухи, в другом, наоборот, возрастание количества осадков, необычно теплые зимы и т. д.
Из непостоянных компонентов атмосферного воздуха экологически для растений наиболее неблагоприятны (как для человека и животных) промышленные газы - диоксид серы, фтор, фтористый водород, хлориды, диоксид азота, аммиак и др. Высокая ранимость "воздушными ядами" растительных организмов объясняется отсутствием у них специальных адаптации к упомянутому, сравнительно недавно возникшему фактору. Относительная устойчивость некоторых растений к промышленным газам связана с их преадаптацией, т. е. наличием тех или иных особенностей, оказавшихся полезными в новых условиях. Так, лиственные деревья легче, чем хвойные, переносят загрязнение воздуха, что объясняется ежегодной листопадностью первых, дающей им возможность регулярно выводить ядовитые вещества с опадом. Однако и у лиственных растений при неблагоприятном газовом составе атмосферы нарушается ритм сезонного развития: задерживается распускание почек, значительно раньше времени наступает листопад.
Из хвойных наименее чувствительны к загрязнению атмосферы туя, некоторые виды можжевельника и ели, особенно сизолистная форма ели колючей (Picea pungens f. glauca), так называемая "голубая ель". Толстостенная эпидерма хвоинок упомянутой формы ели покрыта мощной кутикулой и слоем воска. Вещество кутикулы не растворяется даже в серной кислоте! Не случайно именно указанные древесные растения наряду с лиственницей сибирской успешно используют при озеленении промышленных населенных пунктов. Причем лиственница с ее тонкой и нежной хвоей, в отличие от туи (Thuja), можжевельника и ели высокочувствительна к загрязнению воздуха. Высокая газоустойчивость лиственницы объясняется ее листопадностью. Таким образом, устойчивость растений к загазованности воздуха, как и к другим неблагоприятным экологическим факто
Дата добавления: 2017-02-20; просмотров: 1670;