Роль среднего и заднего мозга в регуляции опорно-двигательного аппарата
1. Существует 5 уровней регуляции двигательной функции.
Первый уровень регуляции формируется на уровне спинного мозга. Здесь в указанной функции участвуют альфа-мотонейроны, гамма-мотонейроны и клетки Реншоу (тормозные нейроны):
1 вариант – от рецепторов кожи в спинной мозг, затем через альфа-мотонейроны на мышечные элементы (кожно-мышечные фазические рефлексы)
2 вариант:
а) от проприорецепторов мышц через альфа-мотонейроны на мышечные элементы (проприоцептивные сухожильные рефлексы)
в) от гамма-мотонейронов через передние корешки спинного мозга на проприорецепторы мышц (интрамуральные мышечные волокна мышечных веретен), а от проприорецепторов к альфа-мотонейронам и через них на мышечные элементы (участие гамма-петли в регуляции мышечного тонуса – проприоцептивные тонические рефлексы спинного мозга)
с) в условиях иннервации мышц-антагонистов на уровне спинного мозга усиление или снижение тонуса флексоров и экстензоров конечности происходит с участием клеток Реншоу (от кожных рецепторов к клетке Реншоу (вставочный нейрон) и через нее к альфа-мотонейрону, иннервирующему мышцу-антагонист.
Таким образом, 1 уровень регуляции двигательной функции предполагает участие: 1 – альфа-мотонейронов
2 – гамма-мотонейронов
LE FMfZjTrt5sg1rwOeHC4mk5CGq1oxd6ouK+7BPU+ex6vlNTNVO4kOyp3p9Y1tJ6dh9j63GZ7JzOms 2DDc8NrSjWse1GifJP+ObO9D1v3DOf4JAAD//wMAUEsDBBQABgAIAAAAIQBl/Z6U4QAAAAsBAAAP AAAAZHJzL2Rvd25yZXYueG1sTI/BToNAEIbvJr7DZky82aUNLYgsjRo9Nca2+gDLMgKRnSXsFqhP 73iqx5n58s/359vZdmLEwbeOFCwXEQgk46qWagWfH693KQgfNFW6c4QKzuhhW1xf5Tqr3EQHHI+h FhxCPtMKmhD6TEpvGrTaL1yPxLcvN1gdeBxqWQ164nDbyVUUbaTVLfGHRvf43KD5Pp6sgrd9PW72 T+59Mu3ubEIof14OO6Vub+bHBxAB53CB4U+f1aFgp9KdqPKiUxBH6ZpRBatlEoNgIknSexAlb9Zx CrLI5f8OxS8AAAD//wMAUEsBAi0AFAAGAAgAAAAhALaDOJL+AAAA4QEAABMAAAAAAAAAAAAAAAAA AAAAAFtDb250ZW50X1R5cGVzXS54bWxQSwECLQAUAAYACAAAACEAOP0h/9YAAACUAQAACwAAAAAA AAAAAAAAAAAvAQAAX3JlbHMvLnJlbHNQSwECLQAUAAYACAAAACEAJi2N6+MCAACABQAADgAAAAAA AAAAAAAAAAAuAgAAZHJzL2Uyb0RvYy54bWxQSwECLQAUAAYACAAAACEAZf2elOEAAAALAQAADwAA AAAAAAAAAAAAAAA9BQAAZHJzL2Rvd25yZXYueG1sUEsFBgAAAAAEAAQA8wAAAEsGAAAAAA== " adj="1747"/> 3 – тормозных нейронов – клеток Реншоу
Вышерасположенные уровни регуляции могут воздействовать на основные звенья 1 уровня и регулировать опорно-двигательный аппарат через воздействие на альфа-мотонейроны, гамма-мотонейроны и клетки Реншоу.
Второй уровень регуляции –варолиев мост и продолговатый мозг
задний мозг
Третий уровень регуляции – средний мозг
Четвертый уровень регуляции –промежуточный мозг
Пятый уровень регуляции –кора больших полушарий и стриопаллидарная
система
Т.о., ИЕРАРХИЯ НЕРВНОЙ РЕГУЛЯЦИИ ДВИГАТЕЛЬНОЙ ФУНКЦИИ:
V IV III II I
Кора БП Промежуточный Средний мозг Варолиев мост и Спинной
мозг продолговатый мозг мозг
II. Второй уровень регуляции – ЗАДНИЙ МОЗГ (продолговатый мозг + варолиев мост)
Второй и последующий уровни регуляции работы опорно-двигательного аппарата являются надсегментарными (сравнить с сегментарным уровнем – спинным мозгом – посегментно замыкающиеся дуги спинальных двигательных рефлексов).
Участие заднего мозга в регуляции двигательной функции прослеживается на таламической лягушке – лягушка, у которой перерезан мозг на уровне зрительных бугров. У животного напряжены экстензоры передних и задних конечностей, организм приподнят над поверхностью опоры – т.е., налицо антигравитационная поза. Это явление имеет эволюционный аспект – амфибия – это первое позвоночное животное, вышедшее на сушу, поэтому создается механизм, обеспечивающий за счет тонуса мышц-разгибателей преодоление силы земного притяжения путем формирования антигравитационной позы. На теплокровном животном этот эксперимент сопровождается развитием состояния децеребрационной ригидности. Это состояние обусловлено повышением тонуса мышц-экстензоров.
Если сделать перерезку выше красного ядра среднего мозга, то ригидность не развивается. Отсюда делаем вывод:
1) Красное ядро тормозит развитие тонуса мышц-разгибателей.
2) Если видоизменить опыт и параллельно перерезке на уровне зрительных бугров сделать перерезку задних корешков спинного мозга, то явление ригидности ослабевает. Следовательно, афферентация от проприорецепторов мышц шеи по путям Бурдаха также обеспечивает развитие этого состояния.
3) Если в условиях децеребрационной ригидности дополнительно произвести разрушение ядер Дейтерса (латеральные вестибулярные ядра), то ригидность исчезает.
4) Такой же эффект (исчезновение ригидности) наблюдается и при разрушении полукружных каналов вестибулярного аппарата.
Таким образом, мы определили 4 влияния, обеспечивающих тонус экстензоров организма – «антигравитационных» мышц организма:
1. Красные ядра (тормозят)
2. Ядра Дейтерса (активируют)
3. Проприорецепторы мышц шеи (афферентация)
4. Вестибулоспинальный аппарат (полукружные каналы) (афферентация)
РАБОТЫ Р. МАГНУСА. Существование определенных нами влияний было показано Р. Магнусом в опытах на кошках, находящихся в состоянии децеребрационной ригидности:
А) участие проприорецепторов шейных мышц в тонических рефлексах продолговатого мозга – у животных с разрушенным вестибулярным аппаратом можно изменить тонус мышц-экстензоров, изменив положение головы по отношению к туловищу. При этом происходит раздражение проприорецепторов мышц шеи, в результате чего происходит рефлекторное перераспределение тонуса в различных мышечных группах: при запрокидывании головы назад повышен тонус мышц-разгибателей передних конечностей и понижается тонус мышц-разгибателей задних конечностей. Противоположные изменения тонуса происходят при агибании головы к грудной клетке.
При повороте головы возникают следующие изменения мышечного тонуса: повышение тонуса разгибателей передней конечности той стороны, куда повернута голова, и снижение тонуса разгибателей конечности противоположной стороны. Эти рефлексы возникают при раздражении проприорецепторов шейных мышц: после перерезки задних корешков шейных сегментов спинного мозга перераспределения тонуса не происходит.
Б) участие лабиринтов вестибулярного аппарата – на шею кошки в состоянии децеребрационной ригидности надевали гипсовую повязку так, что голова по отношению к туловищу занимала постоянное положение (проприорецепторы мышц шеи не могли раздражаться). Затем животное вращали вокруг оси, мысленно проходящей через оба виска. Если животное лежало вверх спиной, тонус мышц-разгибателей уменьшался; если спиной вниз – тонус бы выражен максимально. В этом эксперименте менялось только направление силы тяжести по отношению к телу; отсюда следует, что тонический рефлекс мог начинаться только в тех рецепторах, которые чувствительны к изменению направления земного притяжения, т.е. в рецепторах вестибулярного аппарата.
Аналогичные описанным явления перераспределения тонуса мускулатуры имеют место и у нормального, неоперированного животного. Если над головой кошки держать кусок мяса, то она поднимает голову, при этом передние конечности разгибаются, задние сгибаются и животное садится, принимая позу, удобную для прыжка. Если же перед головой кошки поставить блюдце с молоком, она нагибает голову вниз, что вызывает сгибание передних и небольшое разгибание задних лап, в результате кошкой принимается поза, в которой удобно лакать молоко (отсюда и названия поз Р. Магнусом: кошка, смотрящая на мясо, и кошка, лакающая молоко). Если кошка при шорохе, производимом мышью, поворачивается голову вправо, то благодаря этому движению у нее увеличивается тонус разгибателей правой части тела, дающий лучшую опору, и уменьшается тонус разгибателей на противоположной стороне. Такое перераспределение тонуса благоприятствует дальнейшему движению конечностей именно левой стороны и, следовательно, повороту туловища вправо, в сторону пробегающей мыши.
В заднем мозге расположены ядра черепно-мозговых нервов (с V по VII пары – мост, с VIII по XII – продолговатый мозг), в результате чего этот отдел ствола определяет вегетативное обеспечение рефлексов позы (в продолговатом мозге расположены центры сердечно-сосудистой системы, системы дыхания, функциональной системы пищеварения). Таким образом, задний мозг через распределение тонуса мышц обеспечивает:
1) антигравитационную позу (активация мотонейронов экстензоров);
2) вегетативное обеспечение этой позы (через вегетативные центры, расположенные в нем);
3) устойчивое равновесие при поворотах и наклонах головы.
ОСНОВНЫЕ ФУНКЦИИ ЗАДНЕГО МОЗГА (продолговатый мозг и мост)
| Анализ сенсорной информации | Проводниковая функция | Рефлекторная функция | |
| - первичный центральный анализ информации, поступающий от вестибулорецепторов и слуховых рецепторов | - усиление миотатических спинальных рефлексов (преимущественно мышц-разгибателей) | 1) усиление шейных спинальных рефлексов положения | Интеграция вегетативных рефлексов Сердечно-сосудистых рефлексов Дыхательных рефлексов Пищеваритель-ных рефлексов и др. |
| - первичный центральный анализ информации, поступающей от проприорецепторов и кожных рецепторов головы | 2)формирование вестибулярных рефлексов положения | ||
| - первичный центральный анализ информации, поступающей от многих висцерорецепторов организма | Обеспечение антигравитационной позы тела | Обеспечение относительно устойчивой антигравитационной позы при поворотах и наклонах головы | Рефлексы жизненного обеспечения функций |
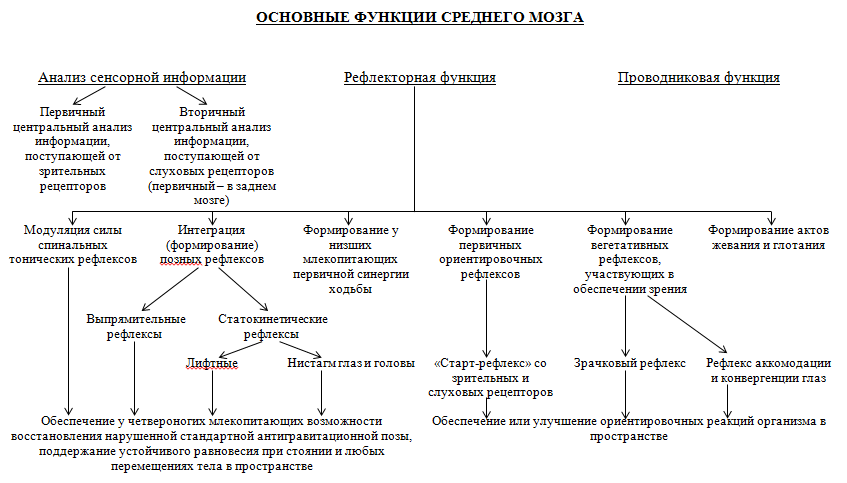
III уровень регуляции – роль среднего мозга (мезенцефальный уровень регуляции)
При перерезке ствола мозга выше красных ядер у мезенцефального животного (магнусовская кошка с сохраненным средним мозгом) отсутствует явление децеребрационной ригидности за счет наличия тонуса экстензоров и флексоров. Поэтому у такого животного проявляются рефлексы выпрямительные, статические:
I фаза:если животное лежит на боку, то оно поднимает голову и устанавливает ее в естественное положение теменем вверх. Это происходит рефлекторно вследствие раздражения вестибулярного аппарата неестественным направлением силы тяжести. При разрушении вестибулярного аппарата поднятие головы все равно осуществляется: раздражаются рецепторы одной половины тела – поэтому происходит рефлекторное выпрямление головы. Если на животное, лежащее на боку, положить сверху доску с небольшим грузом, в этом случае будет иметь место симметричное двустороннее раздражение кожных рецепторов и голова вновь упадет.
II фаза заключается в рефлекторном выпрямлении туловища, которое следует за головой: рефлекс возникает при раздражении проприорецепторов мышц шеи и рецепторов кожи туловища.
В результате: у лежащего животного сначала поднимается голова теменем вверх, в результате раздражаются проприорецепторы мышц шеи и в ответ на это раздражение сокращаются мышцы, выпрямляющие туловище. Таким образом, поднимается голова, а затем, как следствие подъема головы, - туловище, и животное принимает нормальную позу.
Статокинетические рефлексы замыкаются при обязательном участии красных ядер среднего мозга. Проявляются в условиях вращения тела: движение с угловым ускорением (на человеке в кресле Барани) или с линейным ускорением (при стоянии на платформе, которая начинает движение от 0 скорости до максимальной). Проявляются в нистагме головы и глаз: голова сначала медленно вращается в сторону, противоположную вращению тела, насколько это возможно; затем быстрым движением возвращается в нормальное положение относительно туловища. После этого начинается новое медленное вращение в противоположном направлении и вновь быстрый поворот. Глаза реагируют на вращение аналогичным образом: медленным поворотом в сторону, противоположную вращению, и быстрым возвратом в исходное положение.
При движениях тела, когда происходят изменения положения отдельных частей тела, возникает перераспределение мышечного тонуса туловища и конечностей. Если, например, собака поднимает одну лапу, то усиливается тонус в остальных трех лапах, что обеспечивает более устойчивое состояние тела в положении стоя.
Основное влияние от красных ядер осуществляется через руброспинальный путь, который оканчивается посегментно в спинном мозге на экстензорных альфа-мотонейронах (через клетки Реншоу) – тормозное влияние, а через моносинаптическую передачу – на флексорных альфа-мотонейронах – активирующее влияние.
Важнейшей функцией всего ствола является перераспределение тонуса мышц в зависимости от положения тела в простанстве!!!
Работая методом перерезок Р. Магнус на бульбарном и мезенцефальном животных («магнусовские» кошки) всю совокупность разнообразных тонических рефлексов ствола разделил на 2 большие группы:
а) статические рефлексы, которые обусловливают определенное положение тела в пространстве;
б) статокинетические рефлексы, которые вызываются пассивным перемещением тела в пространстве и обеспечивают устойчивое равновесие тела в этих условиях.
Определение: Тоническими рефлексами ствола называются рефлекторные
реакции, которые путем:
1. формирования тонуса мышц;
2. его регуляции;
3. его перераспределения между экстензорами и флексорами
обеспечивают в конечном результате:
1. наличие антигравитационной позы;
2. изменение позы из неестественной в естественную;
3. сохранение позы устойчивого равновесия в условиях пассивного передвижения тела в пространстве.
ТОНИЧЕСКИЕ РЕФЛЕКСЫ СТВОЛА
| Статические | Статокинетические | |||
| Рефлексы положения (позы) или позно-тонические рефлексы | Установочные или выпрямительные рефлексы | Лифтные рефлексы | Нистагм глаз и головы | |
| Функция: | Обеспечивают определенное положение или позу | Обеспечивает возвращение тела из неестественного положения в нормальное | Обеспечивают устойчивое равновесие при любых перемещениях тела в пространстве (с угловым или линейным ускорением или при изменении силы тяжести) или относительном перемещении отдельных его частей | |
| Центры: | Продолговатый мозг (ядра Дейтерса) | Средний мозг (красные ядра) | ||
| Афферентация: | Рецепторы вестибулярного аппарата, проприорецепторы мышц шеи | Рецепторы вестибулярного аппарата, проприорецепторы мышц шеи, экстерорецепторы кожной поверхности тела | ||
| Конечный полезный результат: | Антигравитационная поза при поворотах и наклонах головы | Перераспределение тонуса мышц экстензоров и флексоров и обеспечение позы устойчивого равновесия при пассивном передвижении тела в пространстве |
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ СТВОЛА
В центральной части ствола анатомически выделено образование, состоящее из диффузных скоплений клеток различных типов и размеров, которые густо переплетаются множеством волокон, идущих в разных направлениях. Анатомически описана О. Дейтерсом (2я половина XIX в.) и названа сетчатой или ретикулярной формацией. Подробное ее описание дали В.М. Бехтерев и Рамони-Кахал (близкие по структуре ядра имеются и в таламусе – неспецифические ядра таламуса; волокна от них идут к коре и образуют неспецифические таламокортикальные пути).
Установлено, что ретикулярная формация имеет большое значение в регуляции возбудимости и тонуса всех отделов центральной нервной системы:
1.По нисходящим ретикуло-спинальным путям (медиальному и латеральному) РФ оказывает как активирующее (через γ-мотонейроны), так и тормозящее (через клетки Реншоу) влияние на рефлекторную деятельность спинного мозга.
2.По восходящим ретикулокортикальным путям РФ оказывает активирующее влияние на кору больших полушарий: импульсы от РФ и неспецифических ядер таламуса поддерживают бодрствующее состояние коры (механизм – фоновая деполяризация нейронов коры, влияния неспецифические и специфические).
| Ретикулярная формация ствола | ||
| Основные афферентные связи | ||
| Коллатерали волокон всех восходящих путей ЦНС | Афферентные пути от коры, мозжечка, двигательных ядер ствола | |
| В результате формируется постоянный тонус РФ (более выраженный в ее активирующих структурах) |
| СОСТАВНЫЕ ЧАСТИ РЕТИКУЛЯРНОЙ ФОРМАЦИИ | ||||||||
| Восходящая ретикулярная формация формирует: | Нисходящая ретикулярная формация обладает: | |||||||
| Ретикуло-кортикальные пути | Ретикуло-таламические и ретикуло-гипоталамические пути | Двигательные центры | Вегетативные центры | |||||
| Активирую-щий | Гипноген-ный | Активи-рующий | Тормоз-ный | Специфи-ческие ретикуло-спиналь-ные пути | - Дыха-тельный; - Сосудо-двигатель-ный и др. | |||
| Активирующее и гипногенное влияние на корковые нейроны | Передача сенсорной информации от различных рецепторов организма | Активирующее или тормозное влияние на все спинальные мотонейроны | Активация флексорных и торможение экстензор-ных МН спинного мозга (к осевым мышцам тела) | |||||
| Поддержание должного уровня активности корковых или спинальных нейронов при бодрствовании организма, участие в организации сна | Участие в обеспечении различных двигательных реакций организма | |||||||
РЕТИКУЛО-СПИНАЛЬНЫЕ ВЗАИМООТНОШЕНИЯ
Механизм влияния ретикулярной формации на мышечный тонус стал известен благодаря работам Р. Гранита, который показал, что РФ изменяет активность гамма-мотонейронов спинного мозга. Эти нейроны иннервируют мышечные волокна периферических частей мышечных веретен. Свое название они получили потому, что их аксоны, гамма-эфференты, представляют собой тонкие волокна, относящиеся к типу Аγ, обладающие значительно меньшей скоростью проведения, чем моторные волокна скелетных мышц. Гамма-эфференты, вызывая сокращение мышечных элементов веретен, обусловливает их натяжение и, как следствие этого, усиление афферентной импульсации от рецепторов ядерной сумки веретен. Афферентные импульсы от веретен, постоянно поступая к спинному мозгу, возбуждают альфа-мотонейроны, что и является причиной тонуса мышц. В свою очередь, поток афферентных импульсов от веретен регулируется гамма-мотонейронами.
Ретикуло-спинальные влияния могут быть диффузными, т.е. оказывать влияние на многие сегменты спинного мозга. Это сказывается на тонусе мышц тела и конечностей во время сна.
Ретикуло-спинальные влияния могут быть специфическими, т.е. оказывать влияние на тонус определенной группы мышц.
РЕТИКУЛО-КОРТИКАЛЬНЫЕ ВЗАИМООТНОШЕНИЯ
Путем введения с помощью стереотаксической аппаратуры микроэлектродов в различные участки мозгового ствола Г. Мэгун и Г. Моруцци в 1949 г. обнаружили, что раздражение ретикулярной формации вызывает характерные для пробуждения и состояния естественного бодрствования изменения электрической активности мозговой коры. Аналогичную реакцию можно вызвать раздражением так называемых неспецифических ядер таламуса и дорсального отдела гипоталамуса. При разрушении ретикулярной формации верхних отделов мозгового ствола животное впадает в состояние глубокого сна, хотя афферентные импульсы по специфическим проводящим путям продолжают поступать в сенсорные области больших полушарий. Это доказывает тот факт, что нормальная функция коры больших полушарий и базальных ганглиев в значительной мере зависит от тонизирующих, активирующих влияний РФ мозгового ствола и неспецифических ядер таламуса. В свою очередь, деятельность восходящей активирующей РФ поддерживается от всех рецепторов организма, так как от афферентных волокон, несущих импульсы к таламусу, отходят многочисленные коллатерали к РФ, кроме того, РФ получает импульсы от мозжечка, базальных ганглиев переднего мозга, лимбической системы и коры больших полушарий. Механизм активирующих влияний РФ на кору БП – фоновая деполяризация нейронов коры больших полушарий.
| <== предыдущая лекция | | | следующая лекция ==> |
| Особенности оценки надежности восстанавливаемых изделий | | | Общая характеристика пожаров |
Дата добавления: 2017-10-09; просмотров: 1419;