Пути использования кислорода в клетке
В настоящее время выделено 4 основные пути использования кислорода в организме:
1. Оксидазный путь - окислительное фосфорилирование. Протекает в митохондриях, является основным источником АТФ в аэробных тканях. Потребляет 90% кислорода.
2. Монооксигеназный путь. Обеспечивает включение 1 атома кислорода в молекулу субстрата. Используется для синтеза новых веществ (стероидные гормоны), обезвреживания ксенобиотиков и токсических продуктов обмена в митохондриях и ЭПР.
3. Диоксигеназный путь. Обеспечивает включение молекулы кислорода в молекулу субстрата. Используется для деградации АК и синтеза новых веществ.
4. Пероксидазный и радикальный пути. Кислород участвует в образовании перекисей и активных радикалов, которые необходимы в пероксисомах для внутриклеточного пищеварения, разрушения макрофагами бактерий, вирусов, регуляции метаболизма и т.д. Перекиси и активные кислородные радикалы оказывают также повреждающее воздействие на структуры клеток и тканей, активируя ПОЛ. Разрушение перекисей и инактивация свободных радикалов осуществляется с помощью ферментативной и неферментативной антиокидантной системы.
В соответствие с путями потребления кислорода и катализируемыми реакциями, все оксидоредуктазы разделены на 5 групп:
1).Оксидазы удаляют Н из субстрата (S), используя в качестве акцептора Н только О2, продуктом реакции является вода. Все оксидазы содержат медь. Например, цитохромоксидаза, последний фермент дыхательной цепи.
SН2 + ½О2 → S + Н2О.
2). Аэробные дегидрогеназы, или ФАД зависимые оксидазы, относятся к металлофлавопротеинам (ФАД, ФМН, Fe, Cu, Mo), находятся в пероксисомах и наружной мембране митоходрий. Они отнимают Н от субстрата, и передают его на О2 с образованием перекиси. Дегидрогеназа L-аминокислот (оксидаза L-аминокислот) в почках осуществляет окислительное дезаминирование. Ксантиндегидрогеназа (ксантиноксидаза) превращает пурины в мочевую кислоту. Моноаминоксидазы (МАО) окисляют гормон адреналин и некоторые биогенные амины. Диаминоксидазы (ДАО) окисляют гистамин и другие диамины и полиамины. Образующаяся перекись бактерицидное действие.
SН2 + О2 → S + Н2О2
3). Анаэробные дегидрогеназы многочисленная группа, содержат в качестве коферментов НАДН2, НАДФН2, ФАД, ФМН, цитохромы. Они удаляют Н из субстрата, не используя в качестве акцептора протона О2. Анаэробные дегидрогеназы выполняют две главные функции: перенос Н с одного субстрата на другой в сопряженной ОВР и транспорт электронов в дыхательной цепи. Примеры ферментов: изоцитрат ДГ (НАДН2), сукцинат ДГ (ФАДН2), цитохромы в, с1, с, а и а3.
SН2 + R → S + RН2
4). Оксигеназы. Включают кислород в молекулу субстрата. Работают в составе мультиферментного комплекса, содержащего ФАД зависимую ДГ, Fe2S2-белок, цитохромы Р450 или В5. Этот комплекс встроен в мембрану ЭПР или внутреннюю мембрану митохондрий.
а) Монооксигеназы. Обеспечивают включение 1 атома кислорода в молекулу субстрата. В качестве восстановителей (R) используется НАДФН2, НАДН2 и аскорбат. Участвуют в синтезе стероидных гормонов, обезвреживании ксенобиотиков. Фен-4-монооксигеназа окисляет фен в тир. Пролингидроксилаза (аскорбат) гидроксилирует пролин в проколлагене.
SН + О2 + RH2 → SOH + Н2О + R
б) Диоксигеназы. Обеспечивают включение молекулы кислорода в молекулу субстрата, окисляют циклические соединения с разрывом цикла. Например, триптофаноксигеназа, превращает триптофан в фенилкенуренин.
S + О2 → SO2
5).Гидроксипероксидазы разрушаютперекиси водорода и органические перекиси. Ферментыпероксидаза и каталаза. Пероксидазы, содержат протогем, восстанавливают перекись до воды за счет хинонов, аскорбата или цитохрома с. Глутатионперосидаза, содержит селен, восстанавливает перекись до воды за счет глутатиона.
Н2О2 + RH2 → 2Н2О + R
Каталаза, гемопротеин, использует одну Н2О2 как донор е-, а вторую Н2О2 как акцептор е-, в результате образуется вода и молекула кислорода. Каталазы много в крови, костном мозге, слизистых оболочках, почках и печени.
2Н2О2 → 2Н2О + О2.
Цикл Кребса
ЦТК является процессом окисления АцетилКоА - универсального продукта катаболизма углеводов, жиров и белков. ЦТК протекает в митохондриях с участием 8 ферментов, которые локализованы в матриксе в свободном состоянии, или на внутренней поверхности внутренней мембраны. В ЦТК участвуют 5 витаминов В1, В2, РР, пантотеновая кислота и липоевая кислота в виде коферментов тиаминпирофосфата, ФАД, НАД+, КоА и липоата.
Основной функции ЦТК является образование водородных эквивалентов, которые в цепи окислительного фосфорилирования обеспечивают синтез макроэргических соединений.
Кроме того, ЦТК выполняет ведущую роль в процессах глюконеогенеза, переаминирования, дезаминирования АК, липогенеза и синтеза гема. Интегрирует все виды обмена веществ.
Регуляция ЦТК. Осуществляется с участием 4 регуляторных ферментов: цитратсинтазы, изоцитрат ДГ, α-КГ ДГ и СДГ. ЦТК ингибируется в основном НАДН2 и АТФ, которые являются продуктами ЦТК и цепи окислительного фосфорилирования. Активируют ЦТК в основном НАД+ и АДФ.
Реакции ЦТК
1). Цитратсинтаза локализуется в матриксе митохондрий, ее активируют ЩУК, НАД+; ингибируют АТФ, НАДН2, Сукцинил-КоА, цитрат.
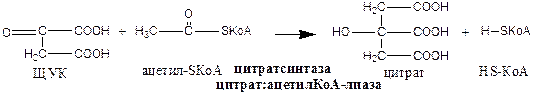
2). Аконитаза локализуется в матриксе митохондрий.
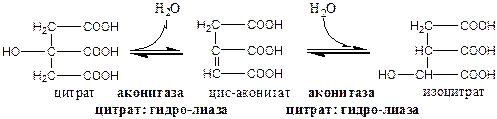
3). Окислительно-восстановительная реакция, самая медленная в ЦТК.
Изоцитратдегидрогеназа локализуется в матриксе митохондрий, ее активируют АМФ, Са2+, АДФ, НАД+; ингибируют АТФ, НАДН2.
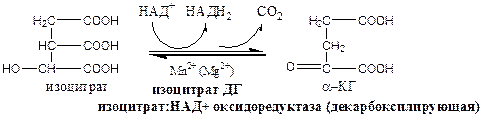
4). Окислительно-восстановительная реакция.
α-кетоглутаратдегидрогеназный комплекс состоит из 3 ферментов и содержит 5 коферментов: тиаминдифосфат, кофермент А, липоевая кислота, НАД+, ФАД.
α-КГ ДГ активируется Са2+, ингибируется сукцинил-КоА, АТФ, НАДН2.
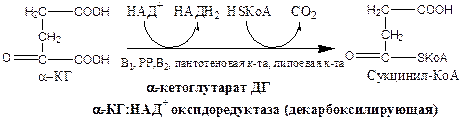
5). Реакция субстратного фосфорилирования
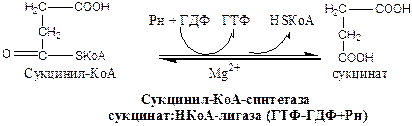
6). Окислительно-восстановительная реакция.
Сукцинатдегидрогеназа, флавопротеин содержащий Fe2S2, связана с внутренней мембраной митохондрии. СДГ ингибирует ЩУК и Сукцинил-КоА,
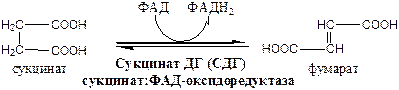
7). Фумараза локализуется в матриксе митохондрий.
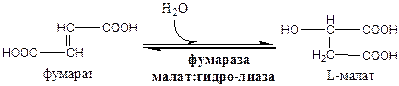
8). Окислительно-восстановительная реакция.
Малат ДГ локализуется в матриксе митохондрий.
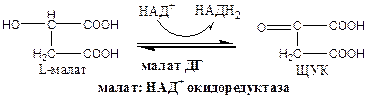
Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь.
Дата добавления: 2017-09-19; просмотров: 735;