Общая характеристика
Согласно представлениям, сложившимся в рамках классической (формальной) генетики, геномы различных организмов характеризуются значительной стабильностью и подвержены лишь очень медленным эволюционным преобразованиям. Эти представления существенно изменились в связи с открытием транспозирующихся (мигрирующих) генетических элементов(фрагментов ДНК), которые могут перемещаться как в пределах одного генома, так и из одного генома в другой. Способность к транспозиции закодирована в нуклеотидных последовательностях самих мигрирующих фрагментов ДНК и связана с возможностью их включения (интеграции) в различные места (сайты) хромосом и внехромосомных молекул ДНК и
выхода («вырезания») из этих сайтов. Такие мобильные структуры были обнаружены в геномах как прокариот, так и эукариот.
Транспозирующиеся элементы первоначально были открыты при обнаружении вставок (инсерций) нового материала в пределах бактериальных оперонов. Такие вставки локализовались внутри гена и предотвращали его транскрипцию и (или) трансляцию. Позднее было установлено, что транспозирующиеся элементы могут непосредственно влиять на геном хозяина, поскольку способны внедрятся не только внутрь генов, но и в близко прилегающие участки, нарушая генную структуру или подчиняя экспрессию этих генов новым регуляторным элементам. Рекомбинация между двумя копиями одного и того же мобильного элемента может приводить к делециям, дупликациям, амплификациям, инверсиям и транспозициям сегментов генома. Мобильные элементы вызывают многие хромосомные мутации и обуславливают их высокую частоту.
У бактерий транспозирующиеся генетические элементы представлены сегментами ДНК двух типов - инсерционными (вставочными) последовательностями или 18-элементами и транспозонами.Те и другие могут перемещаться в границах одной генетической структуры (бактериальной хромосомы, плазмиды) либо из одной структуры в другую например, из хромосомы в плазмиду, из плазмиды в хромосому, из оной плазмиды в другую плазмиду, из плазмиды бактерии одного вида в плазмиду либо хромосому бактерии другого вида и т.д.).
Элементы
Первая группа транспозирующихся генетических элементов получила название инсерционных последовательностей.Каждый тип инсерционной последовательности обозначается префиксом 18,далее следует номер, соответствующий именно этому типу, например, 181, 182, 183 и т.д. 18-элементы были открыты Старлинжером, Сердлером и Шапиро в начале 70-х годов. Они являются нормальными компонентами бактериальной хромосомы и плазмид. В частности, один стандартный штамм кишечной палочки
содержит 5-10 копий 181, 4-8 копий 182, 10-12 копий 1810 ряд других. Для обозначения инсерции в определенном сайте используется двоеточие; так, обозначение лямбда:: 181 показывает, что 181-элемент внедрен в геном фага лямбда.

|
Ген, кодирующий транспозазу-белок, необходимый для перемещения
Рис. 1 18 - элемент кишечной палочки
Строение типичного 18-элемента представлено на рис. 1. 18-элемен-ты - это небольшие участки ДНК, размеры которых обычно варьируют в пределах порядка от 750 до 1550 пар нуклеотидов. На концах (на флангах) этих участков имеются одинаковые либо очень сходные короткие нуклеотидные последовательности (20-40 пар нуклеотидов), расположенные в обратном порядке (инвертированные повторы или 1К-последовательности). Считают, что ^-последовательности играют важную роль в процессах перемещения транспозирующихся элементов. В центральной части все 18-элементы содержат ген, кодирующий синтез белка, названного транспозазой.
В процессе перемещения 18-элементов происходит точное копирование уже встроенного элемента, затем старая копия остается на месте, а вновь синтезированная внедряется в новый сайт. Репликация новой копии происходит с использованием энзимов репликационной машины клетки-хозяина. Процесс транспозиции осуществляется с использованием транспозазы, которая распознает 1К-последовательности, где и инициируется транспозиция.
Внедрение 18-элемента в новый локус сопровождается дупликацией(прямой повтор) последовательностей из нескольких нуклеотидов (5-9 п.н.) данного локуса. Число дуплицированных нуклеотидов специфично для каждого 18-элемента.
Разные 18-элементы являются типичными обитателями бактерий и обычно выявляются в количествах, превышающих одну копию на клетку. Наличие идентичных 18-элементов в разных бактериальных структурах (хромосомах, плазмидах) создает основу для их взаимодействия по механизму сайт-специфической рекомбинации, что может приводить к объединению (коинтеграции) таких структур, либо к генетическому обмену между ними. В отличие от генетической рекомбинации общего типа, требующей наличия значительных областей гомологии ДНК у двух взаимодействующих родственных структур, сайт-специфическая рекомбинация основана на том, что соответствующий фермент (транспозаза) узнает небольшие специфические последовательности в двух местах (сайтах) одной, либо разных молекул ДНК, где и происходит процесс рекомбинации. Следовательно, такая рекомбинация может приводить к генетическому обмену между неродственными структурами (хромосомами, плазмидами).
В целом, 18-элементы представляют собой простейший класс транспозонов; их генетические функции связаны только со способностью к транспозиции.
Транспозоны
Транспозонами (Тп-элементами) называют сегменты ДНК, обладающие такими же свойствами, что и 18-элементы, но содержащие также, гены, не имеющие непосредственного отношения к транспозиции
л 4
' Ь
(гены устойчивости к антибиотикам, гены токсинов, гены дополнительных ферментов клеточного метаболизма и т.д.). В общем, для транспозонов характерны те же гены, которые имеются в плазмидах. Более того, нередко присутствие в составе плазмиды того или иного гена обусловлено наличием в последовательности плазмидной ДНК соответствующего транспозона.
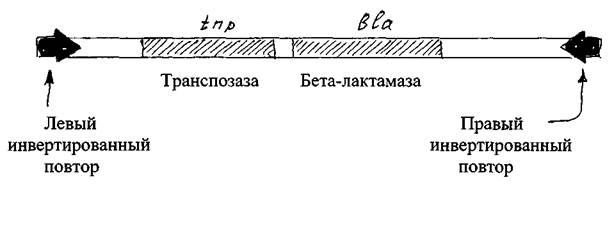
Известны два типа транспозонов прокариот: простые и сложные. Простые транспозоныимеют центральный район, содержащий гены, например, ген синтеза бета-лактамазы, обуславливающей устойчивость к бета-лактамным антибиотикам, и ген, кодирующий синтез транспозазы. Подобно 18-элементам, простые транспозоны всегда имеют на своих концах инвертированные повторы, необходимые для транспозиций (рис. 2).
|
А
Рис. 2 Структура простого транспозона ТпЗ
Сложные транспозонытакже имеют центральный район, содержащий гены. На обоих концах транспозона расположены 18-элементы, которые в пределах одного транспозона принадлежат к одному типу и называются «левый» и «правый». Таким образом, центральный район сложного транспозона фланктрован инвертированными18-элементами, которые, в свою очередь, имеют свои собственные терминальные инвертированные повторы (рис. 3).
^
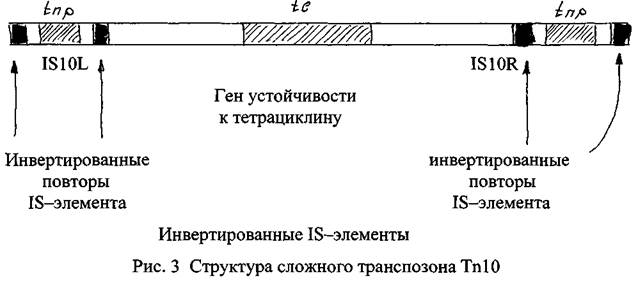
Другое важное отличие сложных транспозонов от простых заключается в том, в большинстве случаев в центральном районе сложных транспозонов отсутствует ген,кодирующий синтез транспозазы, а продукция данного фермента осуществляется с помощью генов, дислоцированных в составе фланкирующих транспозон 18-элементов.
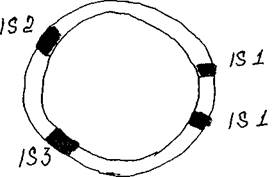
Рис. 4 Образование транспозона близкорасположенными 18-элементами Важно отметить, что часто два близкорасположенных 18-элемента способны перемещаться вместе, одновременно перенося заключенный между ними сегмент ДНК. Таким образом, транспозон могут образовать два расположенных рядом 18-элемента (рис. 4).
Специфические концевые повторы 18-элементов и транспозонов играют ключевую роль в процессах транспозиции, поскольку они распознаются ферментами, осуществляющими сайт-специфическую рекомбинацию. При транспозиции транспозонов работает механизм, как правило, не связанный с процессом полного «вырезания» всей структуры из одного сайта и её перемещения в другой сайт. При этом происходит удвоение (репликация) генетического элемента, одна копия которого остается в исходном сайте, тогда как другая перемещается в новый сайт-мишень. Иными словами, такая транспозиция состоит в увеличении числа копий соответствующего транспозона.
Дата добавления: 2017-09-19; просмотров: 1152;