Основные черты популяции как генетического резерва вида
Как следует из определения, для существования популяциинеобходимы три условия:
– панмиксия (т.е. возможность свободного скрещивания особи с любой другой этого же вида);
– отсутствие изоляционных барьеров, нарушающих панмиксию;
– длительное существование на одной территории.
Панмиксиязаключается в случайном скрещивании родительских пар. Степень панмиксии внутри популяции должна быть выше, чем между популяциями. Эта разница в степени влияния фактора изоляции между особями и определяет обособление групп особей, именуемых популяциями. Однако внутри популяции неизбежно выделяются особи, скрещивания между которыми носит избирательный характер. Причины такой избирательности в лесных популяциях могут быть как чисто механические, так и генетические, Например, местоположение дерева может лимитировать в большей или меньшей степени как разлет, так и попадание на женские цветки пыльцы с тех или иных деревьев. Возможна также конкуренция попавших на рыльце цветка пыльцевых зерен, Причина такой неравноценной возможности у пыльцевых зёрен носит, несомненно, генетический характер, Наконец, генетически обусловлен признак раннего и позднего вступления в фазу цветения у различных деревьев. И все же в популяции в целом влияние факторов, нарушающих панмиксию, не должно быть значительным, так как главная роль панмиксии - обеспечить возможность особям, входящим в популяцию, свободно обмениваться генами, т.е. создать возможно большее число вариаций генотипов предполагаемого видом «рабочего» материала естественному отбору. В популяции, где возможно максимальное перемешивание (комбинация) аллелей, естественный обор оставляет наиболее приспособленные формы организмов. В связи с этим именно панмиксия определяет в первую очередь целостность популяции, как единицы эволюции.
Изоляция нарушает свободную панмиксию внутри популяции, а также ограничивает возможности скрещивания между особями, принадлежащими к разным популяциям. Различают три типа нарушения панмиксии внутри популяции.
Accоpmuвнoе скрещивание – избирательность, при которой особи с одинаковыми генотипами образуют пары чаще, чем при случайном скрещивании. К положительному ассортивному скрещиванию может приводить дизруптивный отбор т.е. отбор генотипов с крайними проявлениями признака. Важнейшим результатом ассортивного скрещивания оказывается изменение частот генотипов.
Инбридинг (инцухт) возможен при повышенной частоте скрещивания между особями, относящимися к одной родственной группе. В этом случае идёт гомоготизация по всем локусам. (рис.)
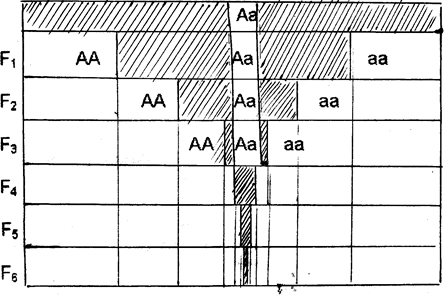
Рис. Динамика соотношения гомо- и гетерозигот при расщеплении по одной паре аллелей при самоопылении:
F1- F6 - поколения инцухта.
Селективное скрещиевание — особи определенных генотипов чаще по сравнению с особями других генотипов вступают в любые скрещивания. В лесных популяциях при таком типе скрещивания отдельные деревья цветут
регулярнее и обильнее, чем другие окружающие их деревья. Селективное скрещивание ведет к изменению в популяциях не только частоты генотипов, но и генов.
Различают два типа межпопуляционной изоляции; территориальную и биологическую.
Изоляция -- явление, обратное миграции. Чтобы популяции генетически дифференцировались, необходима известная степень изоляции. С другой стороны, длительный обмен генами нейтрализует эффект отбора.
При территориальной или географической, изоляции популяции разделены естественным барьером – огромным пространством или горами. Виды, занимающие аридные горные области, имеют сильно разорванные ареалы. Популяции в отдельных горных районов репродуктивно изолированы и тем самым дифференцируются генетически в результате отбора или случайных процессов. Примером разрыва ареалов могут служить островные сосновые боры на южном пределе ареала, берёзовые колки в степной зоне.
Имеются данные, указывающие на очень большой радиус эффективного разлета пыльцы в пределах леса. Для дуба, например, этот радиус составляет несколько сотен метров, а для сосны от 12 – 20 до 200 м., кедр – 250, ель – до 300, ильм – 2000 м. Семена сосен, елей, пихт, клёнов, ясеней, лип остаются в пределах нескольких сотен метров от деревьев с которых они падают.,Семена березы по насту распространяются на несколько сотен метров. У тополей, ив могут лететь на расстояние 0,5 км и более, Семена и дубов, грецких орехов, кедра очень тяжелые и рассеиваться только на несколько метров от дерева. Могут разноситься белками, кедровкой на значительное расстояние. И все же среднее расстояние рассеивания их мало, поскольку большую часть семян белки съедают. Некоторые деревья – можжевельники, вишни, сливы, боярышник, малина поедаются птицами и семена разносятсяна значительное расстояние от родительских деревьев.
Биологическая изоляция у деревьев чаще всего выражается в различных сроках цветения, обусловленных климатическими факторами. Мутации признака раннего и позднего цветения могут привести к репродуктивно
Изолированным популяциям. Но если такие мутации произошли и части популяции начали мигрировать в противоположных направлениях, обор может фиксировать гены раннего цветения в одной части, а гены позднего цветения – в другой. Через некоторое количество поколений дифференциация может стать достаточно большой, чтобы предотвратить скрещивание, если субпопуляции опять сольются. Такой предположительный путь возникновения различий во времени цветения между кленами серебристым (ранний) и красным (поздний). Если сохранить пыльцу, скрестить эти виды искусственно совсем нетрудно. В природе, занимая один и тот же ареал, не скрещивается.
Хозяйственные виды в большинстве своем имеют непрерывное распространение, по крайней мере, на большей части ареалов. Вместе с пыльцой или семенами гены могут мигрировать от одного конца к другому этого непрерывного ареала и таким образом предотвращать генетическую дифференциацию. Однако чаще опыление происходит между близко расположенными деревьями-соседями.
"Соседство" – это полоса длиной 2 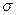
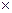
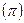 1/2 (или
1/2 (или 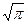 ) (пыльца и семена распространяются одинаково) или
) (пыльца и семена распространяются одинаково) или 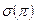 1/2 (распространяется только пыльца).
1/2 (распространяется только пыльца).
Райт установил, что большинство растущих в умеренной зоне древесных видов, имеют средние величины «соседства» 25–200 деревьев, что приводит к небольшой или средней локальной генетической дифференциации в пределах 50 -100км.
Дата добавления: 2017-08-01; просмотров: 1295;