Теория массового отбора и возможность его использования в лесном хозяйстве
При массовом отборе выделяют множество индивидов, наиболее отвечающих задачам селекции, и размножают их совместно. В основе массового отбора лежит оценка фенотипа. Стихийно он применялся человеком с незапамятных времен, затем сознательно использовался при выведении многих высокопродуктивных сортов культурных растений и пород домашних животных. Он не утратил своего значения и в настоящее время, так как он наиболее прост и доступен, а также может нередко давать хорошие результаты.
Однако этот тип отбора имеет и существенные недостатки, обусловленные тем, что по фенотипу трудно однозначно судить о генотипе отбираемых особей, от которого зависит эффективность отбора. Фенотип представляет собой результат взаимодействия генотипа и среды, а на большинство количественных признаков, интересующих селекционера, влияние последней весьма значительно. Кроме того, в фенотипе не проявляются рецессивные гены, находящиеся в гетерозиготном состоянии, но способные проявиться в потомстве. Ввиду этого массовый отбор, как правило, действует очень медленно, а иногда оказывается и вовсе безрезультатным. Последнее характерно главным образом для тех случаев, когда популяция, в которой ведется отбор, гомозиготная или почти гомозиготная по генам, определяющим отбираемый признак, то есть коэффициент его наследуемости близок к нулю.
Хорошей иллюстрацией безуспешности отбора в гомозиготных популяциях, несмотря на наблюдаемую в них фенотипическую изменчивость, могут служить опыты Иоганнсена, проведенные еще в началеXX века. Он изучал изменчивость веса семян фасоли и действие отбора на этот признак. Семена в смеси, собранной с разных растений изучаемого им сорта, характеризовались значительной изменчивостью массы — приблизительно от 0,25 до 0,75 г (средняя масса около 0,5 г). Поскольку нормально фасоль размножается самоопылением, то все семена в этой смеси должны были быть гомозиготными. Однако генотипически они могли быть различными, поскольку происходили от разных родителей. То и другое подтвердилось результатами последующей работы.
Из исходной смеси были отобраны и высажены семена разной массы. Растения, выращенные из наиболее тяжелых семян, дали тоже тяжелые семена (средняя масса около 0,66 г), а растения, выращенные из легких семян, произвели легкие семена (средняя масса около 0,37 г). Т.е. отбор здесь был эффективным. Однако дальнейший отбор самых тяжелых и самых легких семян на протяжении шести поколений в пределах отдельных выделенных таким образом чистых линий оказался совершенно безрезультатным (рис. 2.1), хотя в каждой линии наблюдалась некоторая изменчивость массы семян.
Учитывая значительную гетерогенность лесных древесных пород и трудность применения к ним других видов отбора, массовый отбор широко используется в их селекции. Выбор желательных, наиболее пригодных для селекции типов (как правило, небольшой процент), называют позитивным массовым отбором, а искоренение нежелательных форм — негативным. Последний не следует смешивать с отрицательно действующим отбором, когда из популяции удаляются наследственно ценные компоненты насаждения и сохраняются менее ценные (например, в результате так называемых выборочных или приисковых рубок).
К негативному массовому отбору в лесных популяциях относятся: рубки ухода, когда в первую очередь удаляют наименее ценные, больные и худшие растения; сплошная рубка насаждений, не соответствующих местообитанию для предотвращения опасности распространения непригодных наследственных свойств через пыльцу;
отбор в питомнике, когда удаляют нежелательные типы по ветвлению и устойчивости среди сеянцев и растений.
При позитивном массовом отборе в лесном хозяйстве применяют разные методы:
• отбор лучших географических происхождений, или климатипов;
• отбор лучших эдафотипов в пределах какого-либо географического района;
• отбор лучших (плюсовых) насаждений;
• отбор лучших (плюсовых) деревьев;
• отбор лучших сеянцев в питомниках и семян по крупности и качеству.
Ниже дана краткая характеристика этих видов массового отбора.
2.2.2. Отбор географических происхождений, или климатипов
Изучение насаждений из привозных семян еще в XIX веке показало, что лучшим ростом и качеством отличаются насаждения, созданные из местных семян. Это было замечено при выращивании ели на Тимирязевской даче из семян, привезенных из Германии (Н.С. Нестеров, 1935, цит. поА.В. Альбенскому, 1959). В Воронежской области в культурах сосны, созданных из семян, импортированных из Германии (г. Дармштадт), вырастали большей частью так называемые “пьяные” деревья (М.М. Вересин, 1981). То же явление было отмечено и у других пород, например при выращивании ольхи черной в Германии из семян, привезенных из Бельгии, когда первоначально быстрый рост сменялся его притуплением и заболеванием деревьев в целом.
Исключением из этого правила является южная Швеция, где насаждения ели из семян, привезенных из Средней Европы, растут лучше местных. Это объясняется тем, что после ледникового периода ель не могла проникнуть в южную Швецию с юга через Балтийское море, она постепенно продвигалась туда из северной Швеции через Финляндию, подвергаясь отбору суровым климатом. Наиболее быстрорастущие, но чувствительные к холоду биотипы, вероятно, выпали. В результате этого ели из южной Швеции растут медленнее привозных (Э. Ромедер, Г. Шенбах, 1962). Имеются и другие исключения, когда интродуцированные географические экотипы растут лучше местных (М. Giertych, 1991).
Для изучения влияния географического происхождения или “климатических экотипов” на рост, развитие, устойчивость и качество древостоев уже в начале XIX века были заложены первые опыты Луи-де-Вильмореном во Франции (1823-1832 гг.), М. К. Турским в России (1877-1891 гг.), Кинитцем в Германии (1877-1878 гг.), Цизляром в Австрии (1893-1896 гг.), Энглером в Швейцарии (1890), Шотте в Швеции (1903-1904 гг.), В.Д. Огиевским в Европейской части России (1910-1916 гг.) и др. Проанализировав результаты этих и других опытов, О. Г. Каппер (1946) впервые у нас в стране дал полную картину характера поведения климатических экотипов. Учитывая значение географического происхождения семян для лесного хозяйства, приведем хотя бы в сокращенном виде эти выводы, поскольку на них базировались все последующие исследования географических экотипов, по крайней мере, в нашей стране.
1. Общие:
• Главные лесные породы различного географического происхождения имеют ясно выраженные наследственные свойства по габитусу крон, ветвей и других морфолого-физиологических признаков. Это показали еще опыты Цизляра с силезской и тирольской лиственницами, Энглера и Мюнха с елью и др.
• Переброска семян из одного климатического района в другой должна осуществляться на основании опытов по изучению климатических экотипов.
• Лучший производственный эффект, наибольшую успешность и благонадежность имеют, как правило, посевы в посадках из местных семян.
2. Биологические особенности климатипов:
• Фенология отдельных климатипов имеет свои особенности. Энглер в Швейцарии отмечал, что в 4-5-летнем возрасте разница во времени окончания роста у скандинавских и русских сосен, по сравнению со швейцарскими и южно-германскими, равна двум неделям, а в 6-7-летнем возрасте эта разница доходит до трех недель. В общем, начало вегетации разных климатипов зависит от хода весны. При наступлении сразу высоких температур вегетация начинается почти у всех климатипов (одновременно), тогда как медленное нарастание высоких температур весной вызывает развитие северных и южных форм в разное время. Окончание вегетации происходит у разных климатипов не одновременно. Более северные климатипы заканчивают вегетацию раньше, а южные — позже.
• Отношение к крайностям температуры у отдельных климати-пов различно. Шотте в Швеции отмечал большую повреждаемость морозом, а также большую искривленность стволов от навала снега у южных форм сосен по отношению к местным соснам. В.Д. Огиев-ский наблюдал большую устойчивость северных сосен в условиях Ленинградской области по сравнению с южными формами. Цизляр пришел к заключению, что перенос черешчатого дуба из умеренного климата в более теплый создает морозостойкие культуры. В то же время перенос дубов из теплой области в более умеренный климат усиливает опасность повреждения морозом дубовых культур. В большей части России однолетние сеянцы сосны с наступлением морозов принимают темно-фиолетовый оттенок хвои, что делает их более устойчивыми к морозам, тогда как западноевропейские формы сохраняют ярко-зеленую окраску и нередко побиваются морозами.
• Репродуктивная способность климатипов тоже различна. У северных форм при произрастании их в более южных условиях возраст возмужалости наступает раньше, причем количество шишек у северных форм меньше, чем у местных.
• Северные формы сосны обыкновенной с более выраженным эпидермисом на хвое меньше повреждаются шютте, чем южные формы.
• Семена северной сосны сохраняют свою всхожесть более продолжительное время, чем семена южной.
• Требовательность к свету у климатипов различна. Для отдельных климатипов существует свой оптимум освещения. Так, в условиях Московской области, по Н.С. Нестерову, вологодская сосна менее требовательна к свету, чем владимирская.
• Северные климатипы более устойчивы к навалу снега из-за наличия у них более узких крон, что связано еще и с условиями освещения на севере.
• Дифференциация стволов с возрастом наблюдается в связи с разным отношением к свету, к заморозкам, а также с разной формой кроны и другими свойствами.
3. Изменчивость морфологических признаков у климатипов:
• Форма кроны у деревьев северных климатипов узкая,частопирамидальная, тогда как у южных форм она более широкая и раскидистая.
• Длина хвои и листьев у северных форм более короткая, чем у южных форм.
• Размеры шишек и длина семян у северных климатипов меньше, чем у южных.
• Цвет семян северных климатипов более светлый, чем у южных форм.
• Более быстрый рост соответствует большей величине листьев.
4. Анатомическая изменчивость климатипов:
• Мощность покровных тканей — эпидермиса и гиподермы — в хвое сосны к северу увеличивается. По другим анатомическим признакам определенные тенденции О.Г. Каппер не выявил.
5. Биохимические исследования климатипов:
• Содержание смол увеличивается от северныхк южным климатипам.
• Содержание жиров уменьшается с севера на юг.
6. Анализ роста климатипов:
• Местные растения (в среднем) имеют наибольшие показатели прироста по высоте.
• Дубы, полученные от быстрорастущих материнских деревьев, обнаруживают быстрый рост в высоту, если климат родины и климат места разведения не сильно отличаются один от другого.
• Рост культур (в общем) тем более медленный, чем географически дальше на север, восток или юго-восток отстоят районы заготовок семян от района культур.
• Перенос южных сосен в северные условия, в связи с длительной их вегетацией, приводит к уменьшению процента поздней древесины, а следовательно, и к большей рыхлости и ломкости ветвей и деревьев.
• Коэффициенты формы сосны (q2) увеличиваются от центральных районов на север и северо-восток и уменьшаются на запад, юго-запад и юго-восток.
• Кривизна стволов сосны является признаком наследственного порядка и в то же время может иметь место при нарушении нормальных жизненных условий в результате переноса сосны из одних климатических условий в другие.
Однако О. Г. Каппер не ограничился этими выводами. Он проанализировал методы и условия закладки этих опытов и нашел целый ряд недочетов, которые предложил устранить в последующих работах. В частности, он обращал внимание на необходимость учета не только географических координат, но и экологических условий произрастания насаждений, из которых собираются семена, а также фиксации фенологических форм и даже отдельных деревьев. Он был категорически против составления опытных образцов путем перемешивания семян из разных популяций. Многие из его рекомендаций были учтены при закладке географических культур более позднего времени (М.М. Вересин, 1963; Р.И. Дерюжкин, 1969; А.И. Ирошников, 1977; A.M. Шутяев, 1998 и др.). Накопленный опыт позволил определить оптимальные расстояния по переброске семян тех или иных лесных древесных пород. Усредненные данные, полученные в этих работах, колебались, например, для сосны, по М.М. Вересину (1963), от 300 до 600 км при переброске семян с севера на юг. При переброске их в обратном направлении расстояние уменьшается вдвое. При перемещении семян с востока на запад эти расстояния составляют 500-850 км, в обратном направлении тоже в два раза меньше. В горных районах допускается перемещение семян сверху вниз до 400 м; снизу вверх расстояние уменьшается в два раза.
Более детально опыт по перемещению семян был обобщен в специальном издании “Лесосеменное районирование основных лесообразующих пород в СССР” (1982), на котором следует остановиться более подробно. В основу его, по информации разработчиков, положены следующие данные: результаты исследований внутривидовой изменчивости древесных пород в географических культурах и природных популяциях; материалы по изучению истории расселения видов в межледниковые периоды, численности популяции в разных частях ареала; наличие современных изоляционных барьеров; результаты исследования ареалов других видов растений-эдифи-каторов; особенности плодоношения в многолетнем цикле и качество семян; интегральная оценка природно-климатических факторов; физико-географическое, агроклиматическое, почвенное, геоботаническое, геоморфологическое, лесорастительное и лесохозяйственное районирование.
Кроме того, при его разработке использованы результаты исследований географических культур (семенных потомств географически отдаленных популяций) и рекомендации по переброске семян и лесо-семенному районированию М.К. Турского, А.Н. Соболева, Н.С. Нестерова, В.Д. Огиевского, В.Н. Сукачева, С.А. Самофала, А.П. Тольского, А.И. Колесникова, В.В. Гурского, А.Б. Жукова, П.К. Фальков-ского, Ф.И. Фомина, В.М. Ровского, В.М. Обновленского, О.Г. Кап-пера, М.М. Вересина, А.С. Яблокова, Л.Ф. Правдина, В.П. Тимофеева, С.А. Ростовцева, Е.И. Еньковой и др. Районирование описано раздельно по породам, что связано с различиями в географическом распространении, экологическими особенностями и характером внутривидовой изменчивости главных лесообразующих пород.
За основную единицу лесосеменного районирования принят лесосеменной район, т.е. некоторая территория (в пределах ареала вида) со сравнительно однородными природными условиями и генотипическим составом популяций, с ясно выраженными природными и лесохозяйственными особенностями, которые обусловливают общий характер основных мероприятий по организации лесного семеноводства и использованию однородного по наследственным свойствам семенного материала. Лесосеменной район в ряде случаев разделяется на несколько подрайонов, т.е. на территории, характеризующиеся большей однородностью лесорастительных условий и генотипического состава популяций. Лесосеменные районы расположены на территории одного или нескольких административных подразделений (республика, край, область) или их части.
В пределах ареала вида лесосеменные районы не равноценны между собой по площади, представительству отдельных лесных формаций, перспективам развития семенной базы, лесокультурному фонду данной породы и т.п. Схемы лесосеменного районирования некоторых основных лесообразующих пород нашей страны, взятые из этого документа, приведены на рисунках 2.2 — 2.8.
Для каждого лесосеменного района (подрайона) предусмотрено использование семян из популяций определенного эколого-геогра-фического происхождения. Предпочтение отдается семенам местных и смежных с ними популяций, наиболее адаптированных к природным условиям района. Под местными понимаются семена, собранные непосредственно в пределах лесосеменного района. Семена, заготовленные в других лесосеменных районах, называются инорайонными.
В равнинных областях, отличающихся постепенным изменением биологических признаков деревьев, допускается переброска семян по территории всего лесосеменного района, а также между контактирующими районами и подрайонами. В горных районах, характеризующихся резким изменением условий местопроизрастания в зависимости от высоты местности, перемещение семян рекомендуется лишь в пределах 200-400 м по вертикали от места их заготовки.
В каждом лесосеменном районе (подрайоне) семена предложено собирать раздельно по хозяйственным группам типов леса. В ценных массивах в семенные годы должен обеспечиваться полный сбор семян для последующего использования их в годы неурожая. Для древесных пород с отчетливо выделяющимися фенологическими формами семена заготавливают и используют раздельно по этим формам.
В целом можно отметить, что в цитируемом документе отражен весь опыт, накопленный к тому времени в стране в данном направлении исследований. Указанное лесосеменное районирование было разработано по заданию Государственного комитета СССР по лесному хозяйству (приказ №29 от 6 февраля 1973) отраслевыми и академическими институтами лесного хозяйства и высшими учебными заведениям. Проект сводного лесосеменного районирования скоординирован и подготовлен: Всесоюзным научно-исследовательским институтом лесоводства и механизации лесного хозяйства (Е.П. Проказиным и Б.Н. Куракиным) — по сосне и ели в европейской части СССР; Институтом леса и древесины им. В.Н. Сукачева СО АН СССР (А. И. Ирошниковым) — по сосне и ели в азиатской части СССР, а также по сосне кедровой сибирской, лиственнице и пихте сибирской; Центральным научно-исследовательским институтом лесной генетики и селекции (A.M. Шутяевым) — по дубу;- Тбилисским институтом леса (К.Д. Нацвлишвили) — по пихте кавказской; Украинским научно-исследовательским институтом лесного хозяйства и агролесомелиорации (П. И. Молотковым) — по пихте европейской и буку; Среднеазиатским научно-исследовательским институтом лесного хозяйства (М.Я. Парфеновым) — по саксаулу.
Перечень исследователей, которые участвовали в разработке этого лесосеменного районирования по отдельным породам, насчитывает десятки фамилий. Их список приведен на с. 7 цитируемого сводного документа. Необходимо также отметить активную позицию и существенный вклад в организацию всего этого огромного дела работников тогдашнего Госкомлеса А.И. Новосельцевой и И.Н. Чеботарева.
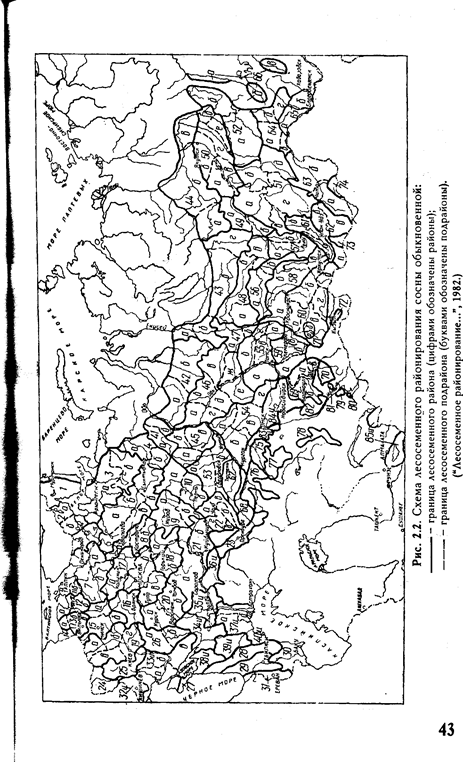
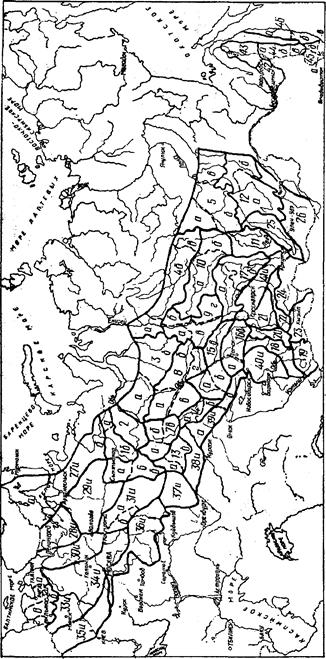
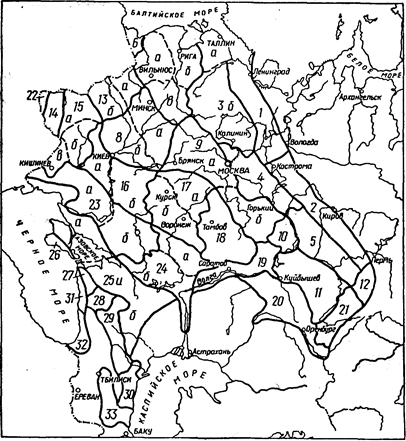
Рис. 2.7. Схема лесосеменного районирования дуба черешчатого:
—— — граница лесосеменного района (цифрами обозначены районы);
•—— — граница лесосеменного подрайона (буквами обозначены подрайоны). ("Лесосеменное районированию...", 1982.)
Учитывать влияние географического происхождения семян на рост насаждений важно не только для автохтонных пород, но в не меньшей степени и для интродуцированных пород. Так, исследование роста различных происхождений сосны скрученной (Pinus contortd) из Канады в Швеции показало, что различные происхождения в разных регионах Швеции показывают разный результат. При этом разница может достигать 60-70 мУга по запасу в 18-летнем возрасте (К. Lindgren, 1993, цит. по Н. Hattemer, F. Bergmann, М. Ziehe, 1993). Следовательно, и при интродукции необходимо прежде всего определять наиболее пригодные части ареала интро-дуцируемой породы, перемещение репродуктивного материала из которых даст наибольший хозяйственный эффект. Этот аспект также был отражен в отечественном лесосеменном районировании.
В рассмотренном выше сводном документе было подготовлено интродукционное районирование основных лесообразующих пород:
а) в европейской части страны по четырем породам: лиственнице — Р.И. Дерюжкиным; ели — Б.Н. Куракиным, A.M. Шутяевым; сосне сибирской кедровой — М.В. Твеленевым, В. И. Некрасовым; сосне обыкновенной — И.Н. Патлаем; б) в азиатской части по интродукции основных лесообразующих пород А.И. Ирошниковым и В.И. Мосиным и др.
Рассмотренный документ был обязательным для всех государственных предприятий и организаций, осуществляющих ведение лесного хозяйства независимо от их ведомственной принадлежности. Все прежние документы утрачивали силу. Однако с течением времени и этот документ потребовал уточнений в части как большей детализации географического происхождения, так и более тщательного учета почвенно-экологических условий.
Дата добавления: 2017-08-01; просмотров: 1171;