МЕХАНИЗМЫ ТОРМОЖЕНИЯ В ЦНС.
Изучив принципы рефлекторной регуляции функций, процессы возникновения и проведения возбуждения, вы поняли, что на все эти процессы можно воздействовать - можно блокировать проведение импульса новокаином, можно увеличить или уменьшить возбудимость ткани, изменив ионный состав окружающей среды, можно воздействовать синаптоактивными веществами. Однако есть механизм, с помощью которого сам организм воздействует на процессы возбуждения и проведения импульса - торможение. Суть этого великолепного достижения эволюции необыкновенно проста: первый вариант - снижение возбудимости клетки вплоть до полной ее невозбудимости, второй вариант - изменение частоты идущих к клетке возбуждающих импульсов вплоть до прекращения проведения. Механизмы торможения - это преподнесенный нам в руки природой еще один путь воздействия на процессы возбуждения и проведения нервного импульса - можно воздействовать на процессы торможения, но для этого их нужно хорошо понять.
С л о в а р ь т е м ы:
Тормозные нейроны
Тормозные медиаторы
ВПСП, ТПСП
Пресинаптическое торможение, Постсинаптическое торможение
Первичное торможение, Вторичное торможение
Торможение Введенского (пессимальное)
Под ТОРМОЖЕНИЕМ понимают самостоятельный нервный активный процесс, вызываемый возбуждением, который проявляется в угнетении или полном выключении другого возбуждения. Торможение обусловлено нервными импульсами, возникающими в особых тормозных нейронах (клетки Рэншоу в спинном мозге, клетки Пуркинье коры мозжечка, звездчатые клетки коры головного мозга и др.).
Впервые возможность влияния на рефлекторные процессы без воздействия на структуры рефлекторной дуги была обнаружена И.М. Сеченовым. В 1863 году И.М. Сеченовым было открыто ЦЕНТРАЛЬНОЕ ТОРМОЖЕНИЕ (СЕЧЕНОВСКОЕ). И.М. Сеченову удалось установить, что сверхпороговое возбуждение центров среднего мозга приводит к торможению периферических, спинальных рефлексов.
Рисунок 23. Схема опыта И.М.Сеченова для демонстрации центрального торможения до и после аппликации на зрительные бугры NaCl.
 Каким же образом можно изменить свойства нейронов так, чтобы они стали невозбудимы? Вспомним, в каких состояниях может находиться нейрон.
Каким же образом можно изменить свойства нейронов так, чтобы они стали невозбудимы? Вспомним, в каких состояниях может находиться нейрон.
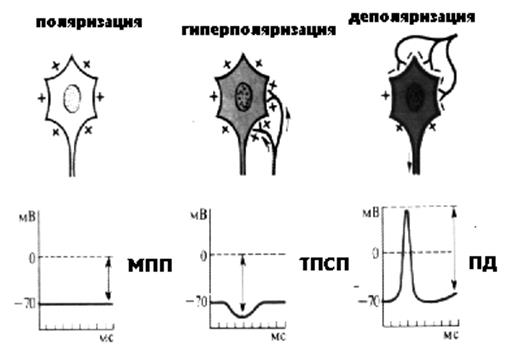
Рисунок 24 Состояния нейрона
На рисунке представлены состояния нейрона: поляризован - обладает мембранным потенциалом покоя, гиперполяризован и деполяризован – потенциал действия. Пока не расшифровывая аббревиатуры «ТПСП», вспомним, что гиперполяризация – это следовой процесс, возникающий при восстановлении МПП после возбуждения. Гиперполяризованный нейрон менее возбудим, потому, что потенциал его мембраны «уходит» от критического уровня деполяризации. Таким образом, снижение возбудимости нейрона может быть результатом длительного возбуждения. Кроме того, возможно простое утомление нейронов, и особенно, синапсов – расходуются запасы АТФ, необходимые для восстановления МПП, запасы медиатора. Следовательно, торможение может быть результатом длительной работы. Такой вариант торможения распространен в ЦНС и получил название вторичного торможения. Один из видов вторичного торможения получил название ПЕССИМАЛЬНОЕ ТОРМОЖЕНИЕ (ТОРМОЖЕНИЕ ВВЕДЕНСКОГО). Такое торможение развивается в результате действия раздражителей с очень высокой частотой. В результате клетка не успевает восстановить свой МПП, остается деполяризованной и, следовательно, невозбудимой.
Нас интересует торможение как самостоятельный процесс. ЦНС обладает механизмами изменения свойств нейрона без предварительного возбуждения. Вспомним результат выделения медиатора пресинаптической мембраной: на постсинаптической мембране происходит деполяризация и возникает локальный ответ. Тормозные нейроны синтезируют другие медиаторы - тормозные. Такими медиаторами могут быть глицин, гамма-амино-масляная кислота (ГАМК). Эти медиаторы, взаимодействуя со своими рецепторами, открывают другие каналы, например калиевые или хлорные. Что же происходит в этом случае с постсинаптической мембраной?

Если вспомнить о том, что ионы калия движутся из клетки, то становится ясно, что при активации калиевых каналов происходит гиперполяризация мембраны и снижение возбудимости нейрона. Тормозной медиатор, как и возбуждающий, активирует рецептор управляемые каналы, однако не натриевые, а калиевые. Эта активация вызывает увеличение выхода ионов калия наружу и ГИПЕРПОЛЯРИЗАЦИЮ мембраны. Изменение потенциала мембраны при гиперполяризации называется тормозной постсинаптический потенциал (ТПСП). Во многих тормозных синапсах аналогичный эффект имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Результатом ТПСП является удаление МПП от критического уровня деполяризации возбуждение становится либо вовсе невозможным, либо для возбуждения требуется значительно большая сила раздражителя. Нервные импульсы, возникающие при возбуждении тормозных нейронов, вызывают гиперполяризацию постсинаптической мембраны и тормозной постсинаптический потенциал (ТПСП), это наиболее распространенный тип торможения в нервной системе, который называется ПОСТСИНАПТИЧЕСКИМ ТОРМОЖЕНИЕМ. Постсинаптическое торможение является первичным торможением – самостоятельный процесс, вызванный возбуждением тормозных нейронов.
Понятно, что в случае деполяризующего медиатора изменение потенциала мембраны будет называться возбуждающий постсинаптический потенциал (ВПСП).
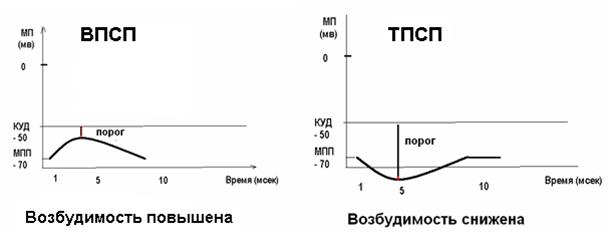
Рисунок 25 Изменение мембранного потенциала при ВПСП и ТПСП
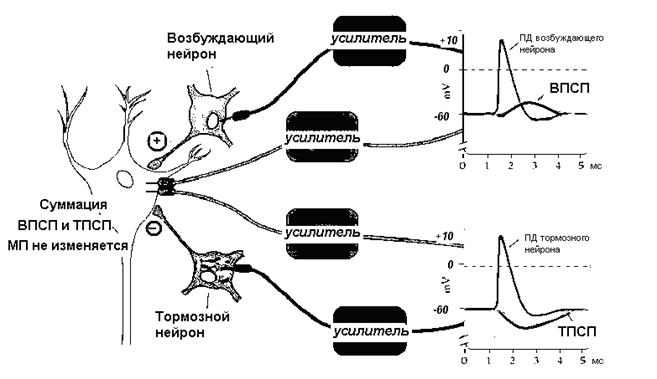
Рисунок 26 Суммация потенциалов на мембране нейрона
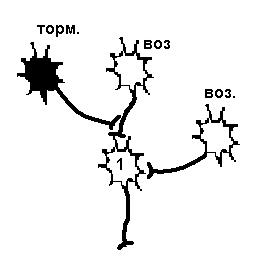
Центральные синапсы отличаются тем, что медиаторы вызывают появление на постсинаптической мембране только локального ответа, (ВПСП или ТПСП). ВПСП обладает всеми свойствами ЛО - зависит от силы раздражителя (количества выделившегося медиатора), суммируется и не передается по аксону. Амплитуда одиночного ВПСП довольно мала - несколько мВ, поэтому для уменьшения заряда мембраны до критического уровня необходима либо одновременная активация нескольких возбуждающих синапсов - ПРОСТРАНСТВЕННАЯ СУММАЦИЯ, либо повышение частоты поступающих импульсов - ВРЕМЕННАЯ СУММАЦИЯ. Если одновременно с ВПСП на мембране нейрона формируется и ТПСП, то эти изменения мембранного потенциала суммируются, следовательно, если величины ВПСП и ТПСП примерно одинаковы, то изменения мембранного потенциала нейрона, на котором суммируются воздействия, не произойдет. На рис.26 представлена схема регистрации мембранного потенциала возбуждающего и тормозного нейронов, которые одновременно воздействуют на один нейрон. Изменений мембранного потенциала нет. На рис 27 слева два возбуждающих нейрона, действуя на нейрон 1 одновременно, могли бы вызвать его возбуждение. Если одновременно на этот нейрон подействует тормозной, то возбуждения не произойдет.
Существует еще один вариант первичного торможения в ЦНС: ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ. Такое торможение возникает в том случае, когда нервный импульс или просто не достигает синаптического окончания аксона, или снижается его частота. Такое торможение осуществляется благодаря аксоаксональным синапсам, образованным терминалями аксонов тормозных интернейронов и аксонами возбуждающих нейронов (рис.27, справа). В этих синапсах торможение определяется действием ГАМК, открывающей хлорные каналы на пресинаптических мембранах (мембрана возбуждающего нейрона). Из - за высокого МПП мембраны аксона (действие электрического, а не концентрационного градиента) ионы хлора выходят через мембрану наружу. Выход ионов вызывает незначительную деполяризацию постсинаптической мембраны, являющейся в данном случае участком мембраны аксона другого нейрона. Деполяризация мембраны аксона ведет к инактивации натриевой проводимости. Возникает блокирование проведения нервного импульса, что уменьшает частоту, или полностью угнетает возбуждающий потенциал, идущий к клетке по возбуждающим нервным окончаниям.
| Постсинаптическое торможение | Пресинаптическое торможение |
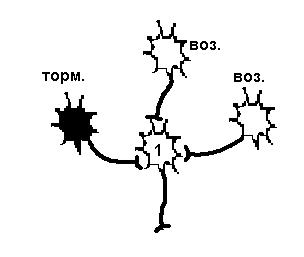
| 
|
| Возбудимость нейрона 1 снижена в результате гиперполяризации мембраны, вызванной действием тормозного нейрона, поэтому два возбуждающих нейрона не могут вызвать ПД. | Возбудимость нейрона 1 не изменена, но возбуждающий импульс от одного из возбуждающих нейронов не доходит, а один возбуждающий не может вызвать ПД. |
Рисунок 27 Первичное торможение
Дата добавления: 2017-08-01; просмотров: 4833;