Гетерозисные скрещивания
Цель таких скрещиваний состоит в получении гибридного посадочного материала, обладающего гибридной мощностью, жизнеспособностью, повышенной продуктивностью или улучшенными качествами у гибридов первого поколения. Гетерозис считается истинным, если наблюдается превосходство гибрида по какому-нибудь признаку над признаком лучшего родителя, и гипотетическим, если превосходство наблюдается над средним значением признака обоих родителей.
А. Густафссон (1951, цит. по Р. Ригер и А. Михаэлис, 1967) под- , разделяет гетерозис по типу проявления на соматический, репродуктивный и приспособительный. Между этими тремя типами существуют переходы. Гетерозис соматический — это более мощное развитие вегетативных органов у гибридных растений; репродуктивный — более мощное развитие репродуктивных органов и повышенная фертильность, приводящие к формированию высокого урожая семян или плодов; приспособительный, или адаптивный, — повышение приспособленности гибридных организмов к изменяющимся условиям среды и их конкурентной способности в борьбе за существование.
Для объяснения причин гетерозиса разработано несколько гипотез. Наиболее распространена гипотеза доминирования, которая основана на представлении о том, что в процессе эволюции гены, благоприятно действующие на организм, становятся доминантными или полудоминантными, в то время как гены, действующие неблагоприятно, становятся рецессивными. Согласно этой гипотезе гетерозис объясняется тремя эффектами действия благоприятных доминантных генов: 1) подавление вредного действия рецессивных аллелей: Аа > аа; 2) аддитивный (суммирующий) эффект неаллель-ных доминантных генов, однонаправленно действующих на определенные количественные признаки, по которым в большинстве случаев и наблюдается гетерозис: А+В + С>А + В,А + Сили В + С >А, В или С (этот эффект присутствует и в случае трансгрессии, о чем было сказано выше); 3) комплементарное взаимодействие ряда не-аллельных доминантных генов: А <=> В > А + В.
Гипотеза сверхдоминирования объясняет гетерозис аллельным взаимодействием генов в гетерозиготном состоянии, вследствие чего АА < Аа > аа. Предполагается, что одинарная доза гена А благоприятнее действует на организм, чем его двойная доза в гомозиготе АА. Кроме того, действие разнонаправленных и независимых аллелей А и а при соединении их в гетерозиготу приобретает характер совместного однонаправленного доминирования; Ах а =* АА', АА < АА'> аа. Одним из доказательств обоснованности теории сверхдоминирования является факт моногибридного гетерозиса, проявляющегося при скрещивании двух гомозиготных линий, различия между которыми состоят только в генах одной аллельной пары: AABBCCDD х aaBBCCDD => AaBBCCDD. При этом АА... < Аа... > аа.... Гипотеза генетического баланса объясняет явление гетерозиса суммарным эффектом разнородных генетических процессов, изменяющих генетический баланс у гетерозиготы в сторону проявления той или иной формы гетерозиса.
Независимо от сущности разных теорий, объясняющих явление гетерозиса, с практической стороны важно иметь в виду, что гетерозис проявляется главным образом в Fv
Н.В. Старова (1980) у древесных пород выделяет три категории гетерозиса: популяционный, групповой и индивидуальный; а в каждой категории — типы гетерозиса: по характеру его проявления (генеративный, соматический, адаптивный) и по характеру взаимодействия генов (доминирование, сверхдоминирование, аддитивные и комплементарные эффекты). Поскольку описание типов было дано ранее, остановимся на характеристике категорий.
Популяционный гетерозис может возникать в результате длительной адаптивной эволюции в панмиктических естественных популяциях в результате удачных рекомбинаций и уравновешивающего естественного отбора, благоприятствующего гетерозиготам. Он может быть получен при географически отдаленной межпопуляционной гибридизации или при гибридизации различных природных изолятов в результате рекомбинации генов. В бывшем СССР были начаты работы по созданию плантаций сосны обыкновенной (И.Н. Патлай, П.И. Молотков), лиственницы (Ф.Д. Авров), березы (А.Я. Любавская) и других пород с использованием межпопуляционных скрещиваний географически отдаленных экотипов и изолятов.
Групповой гетерозис может быть получен при искусственной гибридизации родительских форм с высокой специфической комбинационной способностью, когда гибридная семья в целом по средним и максимальным показателям превосходит обе родительские формы. Примеры такого гетерозиса встречаются у тополей, ив и других пород.
Индивидуальный гетерозис наблюдается при межвидовых или географически отдаленных скрещиваниях, а также при скрещивании растений с различным уровнем плоидности. В этом случае не гибридные семьи в целом, а лишь отдельные экземпляры превосходят родительские формы (С.З. Курдиани). Индивидуальный гетерозис можно использовать в клоновой селекции при вегетативном и апомиктическом размножении гетерозисных форм для создания высокопродуктивных плантационных насаждений. Наиболее часто этот вид гетерозиса применяют при гибридизации тополей, ив, орехов рода Juglans L.
3.2. Методы гибридизации
Методы гибридизации зависят от биологических особенностей вида, характера исходного материала, требований к будущему сорту идр. Выделяют простые и сложные скрещивания, кроме того, существует ряд методов, сочетающих гибридизацию с другими подходами хозяйственного улучшения растений (инбридинг, полиплоидия, мутагенез и др.).
Простые скрещивания. Скрещивания между двумя родительскими формами, производимые однократно, называют простыми:
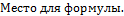
При простых скрещиваниях 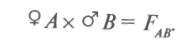 гибриды получаются от объединения наследственности двух родителей. Формообразовательный процесс в гибридных популяциях от простых скрещиваний идет на основе перераспределения наследственного материала, привнесенного в равном количестве одной парой родителей, поэтому простые скрещивания называют также парными. Разновидностью парных скрещиваний являются взаимные или реципрокные, когда в качестве материнского растения выступает растение, бывшее ранее отцовским:
гибриды получаются от объединения наследственности двух родителей. Формообразовательный процесс в гибридных популяциях от простых скрещиваний идет на основе перераспределения наследственного материала, привнесенного в равном количестве одной парой родителей, поэтому простые скрещивания называют также парными. Разновидностью парных скрещиваний являются взаимные или реципрокные, когда в качестве материнского растения выступает растение, бывшее ранее отцовским:
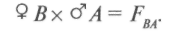
Реципрокные скрещивания применяются:
а) когда наследование какого-либо важного хозяйственно-биологического признака (например, зимостойкости, роста и др.) связано с цитоплазмой; гибриды наследуют это свойство сильнее в том случае, если носитель ценного признака берется в виде материнского растения;
б) когда завязываемость семян зависит от того, в качестве материнской или отцовской берется та или иная форма.
Сложные скрещивания. В сложных скрещиваниях участвуют более двух родительских форм, или гибридное потомство повторно скрещивается с одним из родителей. Они делятся на ступенчатые и возвратные. Ступенчатые скрещивания применяются, когда необходимо соединить в гибридном потомстве наследственность нескольких родительских форм. Их можно представить в виде следующих простейших формул:
1)[(АхВ)хС]х D; 2)[(AxB)x(Cx D)] х Е.
В первом случае гибрид, полученный от скрещивания двух родительских форм А и В, дополнительно скрещивается с формой С, а затем с формой D; в целом здесь объединяется наследственность четырех родительских форм. Во втором случае сначала скрещиваются попарно формы А и В, С и D, а их гибридное потомство скрещивается между собой и с формой Е; в целом здесь объединяется наследственность пяти родительских форм. В обоих случаях скрещивания осуществляются последовательно, ступенчато.
Возвратные скрещивания — это такие скрещивания, при которых гибрид повторно скрещивается с одним из родителей. Их применяют в двух случаях: 1) для преодоления бесплодия гибридов первого поколения при отдаленной гибридизации: (Ах В) х В; 2) для усиления в гибридном потомстве желаемых свойств одного из родителей. В этом случае возвратные скрещивания называют насыщающими. Чаще всего их используют при выведении сортов, устойчивых к болезням или неблагоприятным абиотическим факторам среды. Лучший высокопродуктивный, но неустойчивый к действию какого - либо неблагоприятного фактора сорт берется в качестве отцовской формы, а сорт, обладающий устойчивостью к нему, — в качестве материнской. Например, скрещиваются устойчивый сорт А с быстрорастущим, но мало устойчивым сортом В, тогда мы можем иметь следующую последовательность возвратных скрещиваний, каждое из которых называется беккроссом (табл. 3.2). В результате первого беккросса количество отцовского ядерного материала увеличивается до 75%, после пятого — оно равняется 99,2%, т.е. происходит почти полное поглощение материнской наследственности отцовской, поэтому такие насыщающие скрещивания называются поглотительными.
Использование в гибридизации явления инбридинга позволило разработать инцухт-гетерозисный метод получения гетерозисных растений. При этом методе сначала получают инцухт-линии или инбред -линии (самоопыленные линии), представляющие собой потомство одного перекрестно опыляющегося растения, полученное в результате принудительного самоопыления. Исходное растение, которое принудительно подвергли самоопылению, обозначается символом 1о, первое его инбредное потомство — символом 11, второе — символом 12 и т.д. Затем отбирают инбредные линии, обладающие высокой комбинационной способностью, и скрещивают их между собой. В результате получают гибриды, обладающие повышенным гетерозисом. Такие гибриды называют инцухт-гетерозисными. Прекрасные результаты применения этого метода получены на сельскохозяйственных растениях, в частности у кукурузы. О его применимости для лесных пород указывал еще А.И. Колесников, а в более позднее время — Э. Ромедер и Г. Шенбах (1962).
Таблица 3.2
Схема применения насыщающих скрещиваний в селекции на устойчивость (по Г.В. Гуляеву, А.П. Дубинину, 1974)
| Номер беккросса | Последовательность скрещиваний | Процент насыщения устойчивого сорта свойствами лучшего высокопродуктивного сорта |
| А х В = F1; | ||
| F1*B = F2 | ||
| F2*B = F3 | 87,5 | |
| F3*B=F4 | 93,7 | |
| F4 *B = F5 | 98,4 | |
| F5*B = F6 | 99,2 |
Необходимо отметить, что в зависимости от способа размножения те или иные методы гибридизации и селекции дают более или менее важные практические результаты.
Так, для самоопылителей наибольшее распространение получили методы комбинационной селекции, как простые, так и сложные, особенно метод ступенчатой и отдаленной гибридизации. Среди перспективных методов можно отметить: использование принципов генетически регулируемого гетерозиса; получение индуцированных мутаций при применении радиации и химических мутагенов и использование искусственно получаемых аллополиплоидов (Н.П. Дубинин, Я.Л. Глембоцкий, 1967).
В селекции перекрестников сельскохозяйственных культур исторически прослеживается переход от массового отбора к семейственному отбору и затем к получению самоопыленных линий и использованию гетерозисных гибридов. Здесь, как и при скрещивании самоопылителей, наблюдаются простые и сложные скрещивания. Новыми методами в этом случае являются скрещивание инцухт-ли-ний (межлинейные гибриды) или скрещивания инцухт-линий на сорт (сортолинейные гибриды), при которых проявляется гетерозис.
В селекции вегетативно размножаемых растений наибольшее значение приобрели такие методы селекции, как отдаленная гибридизация, экспериментальное получение полиплоидов и искусственный мутагенез. При всех этих методах важно получить хотя бы одно растение с улучшенными хозяйственными признаками, которое в последующем можно размножить вегетативным путем. Успех здесь зависит в основном от выбора исходного материала и технического осуществления методов.
3.3. Техника гибридизации
Проведение контролируемых скрещиваний лесных деревьев — очень дорогое и долговременное мероприятие. Поэтому прежде чем егопроводить, составляют схему (план) скрещиваний, при разработке которой стараются сделать так, чтобы можно было одновременно достичь нескольких целей. Различают самоопыление, системы скрещиваний с неизвестными отцами (свободное опыление и поликросс) и системы скрещивания с известными отцами. Планы скрещиваний составляют, собственно говоря, только для последней группы скрещиваний. Различают полную схему диаллельных скрещиваний, модифицированную схему диаллельных скрещиваний, мистичные схемы диаллельных скрещиваний, факториальную схему скрещиваний, простые скрещивания (В. Zobel, J. Talbert, 1984, И. Ditlevsen, 1985).
Полная схема диаллельных скрещиваний (рис. 3.1) считается одной из лучших, поскольку включает все возможные варианты скрещивания и дает почти полную информацию о генетических характеристиках изучаемых клонов. Схема может давать информацию об ОКС и СКС и их дисперсиях. Материал также создает лучшую стартовую точку для отбора наилучших индивидуумов или пар клонов, подходящих для создания биклоновых плантаций. Схема, к сожалению, очень трудна для осуществления, особенно с экономической точки зрения. Например, для реализации полной диаллельной схемы для 20 клонов надо провести 400 контролируемых скрещиваний или 380, если исключить самоопыление. Поэтому в практике используют другие схемы (рис. 3.2-3.4).
Модифицированная схема диаллельных скрещиваний представляет собой ограниченную диаллельную схему скрещиваний. В этом случае из полной схемы исключаются реципрокные скрещивания и самоопыления (рис. 3.2), что значительно удешевляет реализацию схемы. Эта схема дает примерно такую же информацию, как и полная схема диаллельных скрещиваний, но ограничения, допущенные в схеме, не гарантируют такой же точности в опытах при определении параметров.
Частичная схема диаллельных скрещиваний может отличаться от полной и модифицированной схемы диаллельных скрещиваний настолько, что один клон не скрещивается со всеми другими клонами (рис. 3.3). Схема менее эффективна, чем схемы полных и модифицированных диаллельных скрещиваний. Однако это компенсируется тем, что большое количество потомства может быть испытано при довольно низких затратах. К сожалению, отсутствие оценок, которые нельзя получить в частичных схемах, значительно усложняют расчеты.
| Матери | Отцы | |||||||||
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X |
Рис. 3.1. Полная схема диаллельных скрещиваний: X — самоопыление
| Матери | Отцы | ||||||||
| X | |||||||||
| X | X | ||||||||
| X | X | X | |||||||
| X | X | X | X | ||||||
| X | X | X | X | X | |||||
| X | X | X | X | X | X | ||||
| X | X | X | X | X | X | X | |||
| X | X | X | X | X | X | X | X | ||
| X | X | X | X | X | X | X | X | X |
Рис. 3.2. Модифицированная схема диаллельных скрещиваний
| Матери | Отцы | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X |
Рис. 3.3. Частичная диаллельная схема Брауна
Факториальная схема скрещивания отличается тем, что в ней все материнские клоны скрещиваются с одними и теми же отцовскими клонами (рис. 3.4). Часто это может быть небольшое число отцовских растений, называемых также общими тестерами. Данная схема может также рассматриваться как разновидность полной диаллельной схемы, включающей все комбинации одной группы матерей и другой группы отцов. Схема очень распространена в США под названием Северо-Каролинская II. Нормально четыре различных отца используются для схемы скрещивания, но, как правило, число отцов в схеме зависит от значения специфических комбинационных эффектов. Так как схема часто включает очень немногое количество отцов и одни и те же клоны не являются одновременно материнскими и отцовскими, то трудно сравнить ОКС каждого родителя. Трудно также отобрать потомство для следующего поколения семенных плантаций, особенно если используется только несколько отцов, являющихся часто уже родственными друг другу. Преимущество этой схемы в простоте ее выполнения и относительной дешевизне; в то же время анализ результатов легче.
| Матери | Отцы | |||
| X | X | X | X | |
| X | X | X | X " | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X |
Рис. 3.4. Факториальная схема скрещивания
Простое скрещивание пар характеризуется тем, что в этом случае каждый клон включен только один раз — как мать или как отец. Эта схема особенно хороша, если целью является создание популяции для отбора особей для новых семенных плантаций или для использования в дальнейшей селекционной работе. Другим преимуществом использования простого скрещивания пар является то, что большое количество клонов может быть испытано при одинаковой схеме, и обычно очень дешево может быть получено потомство на базе контролируемых скрещиваний. С другой стороны, возможности оценки ОКС и дисперсии ОКС и СКС обычно не очень хороши. Выбор соответствующей схемы зависит от цели контролируемых скрещивании. В табл. 3.3 суммированы достоинства и недостатки названных схем.
При планировании скрещиваний следует учитывать ряд особенностей исходного материала, отмеченных С.С. Пятницким (1961):
• для получения крепкого потомства необходимо отбирать для скрещивания здоровые неистощенные экземпляры;
Таблица 3.3
Сравнение схем скрещиваний
(по В. Ditlevsen, 1985)
| Система скрещивания | Оценка ОКС | Отбор плюсовых деревьев | Затраты | Оценка дисперсии ОКС и СКС |
| Свободное опыление | Неплохая | Возможен, но не эффективен | Низкие | Затруднена |
| Поликросс | Очень хорошая | Очень незначителен и только, если дисперсия близкородственного скрещивания ослаблена | Очень низкие | Хорошая оценка дисперсии ОКС |
| Полная диаллельная | Превосходная | Превосходный | Очень высокие | Превосходная |
| Модифицированная диаллельная | Превосходная | Превосходный | Очень высокие | Очень хорошая |
| Частичная диаллельная | Хорошая | Очень хороший | Достаточные | Хорошая оценка дисперсии ОКС. Дисперсия СКС может быть определена, но это трудно с точки зрения обработки данных |
| Факториальная | Хорошая | Только в немногих случаях и тогда, когда дисперсия близкородственного скрещивания низкая | Достаточные | Хорошая |
| Простое скрещи-h.niHc пар | Плохая | Хороший | Очень низкие | Плохая |
• у молодых растений выход гибридных семян и плодов выше, чем у старых;
• при скрещивании с местными видами полученные гибриды больше уклоняются в их сторону;
• при скрещивании культурной формы с дикой уклонение идет в сторону последней;
• при скрещиваниях материалов из географически отдаленных местностей или экологически неоднородных условий местопроизрастания больше вероятности получить хозяйственно ценные гибриды;
• чем более отдаленные в филогенетическом смысле исходные пары, тем труднее проводить скрещивания; при этом результаты, полученные для пар в пределах секции, могут оказаться выше результатов, полученных для более отдаленных пар.
Искусственные скрещивания можно проводить на стоящих деревьях, привитых деревьях, срезанных (укорененных или не укорененных) ветвях, на поваленных деревьях.
При скрещиваниях на стоящих деревьях для подъема в крону используются лестницы, подмостки или специальные подъемники. Работа на стоящих деревьях трудоемка и опасна.
Проще работа на привитых деревьях ввиду возможности осуществить все желательные комбинации независимо от местообитания , исходных особей. Так как привойный материал можно собирать и перевозить даже с одного континента на другой, то участки для скрещивания можно создавать и с географически отдаленными растениями и видами. Прививку ветвей можно осуществлять на подвои, высаженные в сосуды. Известен метод бутылочной прививки, который состоит в соединении больших цветоносных ветвей с такими подвоями. При этом привой до срастания снабжается водой из бутылки. Этот метод используется для березы, ольхи, тополя, вяза.
Скрещивания на срезанных ветвях, помещенных в сосуды с водой, применяются у тополей, ив, ильмовых и некоторых других пород. Ветви нарезают в конце зимы. Чем ближе к естественным срокам цветения заготавливают ветви и чем они крупнее, тем успешнее результаты гибридизации. Женские ветви рекомендуется нарезать длиной 1 м и больше. Скрещивания лучше проводить в помещении с регулируемой температурой и влажностью.
Скрещивания на поваленных (или подваленных) деревьях значительно упрощают работу. Однако недостатком является разовое использование отобранных деревьев и невозможность быстрого повторения наиболее удачных скрещиваний (если это не клонированные особи).
Цветки на отобранных для проведения гибридизации женских особях за 1-2 недели до скрещивания подвергаются изоляции и кастрации во избежание неконтролируемого опыления. При кастрации удаляют мужские генеративные почки, цветки или часть цветков на ветвях, подлежащих изоляции. Обычно кастрация у обоеполых цветков выполняется путем выщипывания тычинок пинцетом. Однополые мужские цветки и генеративные почки удаляются полностью. У двудомных и однодомных растений с однополыми цветками, у которых последние пространственно разобщены, кастрацию не производят.
Для изоляции женских цветков и соцветий применяются изоляторы из различных материалов: синтетическая пленка (полиэтилен, целлофан); пергаментная бумага; бумага, пропитанная водоотталкивающим составом; ряд тканей. Преимущества и недостатки каждого вида материала показаны в табл. 3.4. Предпочтение в практической работе отдается пакетам из пергамента (см. рис. 3.5 на вклейке). При изоляции необходимо следить за тем, чтобы в месте привязки пакета не оставалось щелей для залета пыльцы. Для уплотнения, а также обеспечения воздухообмена используют вату в месте привязки пакета.
Размеры изоляторов зависят от размеров изолируемых ветвей или цветков. Они могут колебаться от 2-3 х 7-10 см до 20-40 х 50-60 см.
Пыльцу, необходимую для гибридизации, заготавливают заблаговременно за несколько дней до цветения (если используется свежесобранная пыльца). Крупные мужские соцветия в почти зрелом возрасте обрывают и помещают в бумажные пакеты или специальные эксикаторы, которые располагают в теплом помещении. После подсыхания пыльники растрескиваются и пыльца собирается в специальные сосуды. Если мужские соцветия мелкие, они нарезаются вместе с мелкими ветвями и раскладываются на бумаге в изолированных помещениях. После выделения пыльцы, ее просеивают для удаления примесей.
При несовпадении сроков цветения родительских форм, их территориальной разобщенности и при дальних перебросках пыльцы прибегают к более ранним срокам ее получения на срезанных ветвях в теплом помещении. Для нормального развития цветочных почек регулярно меняют воду или питательный раствор в сосудах и проводят подновление срезов.
Пыльца быстро теряет всхожесть, поэтому ее необходимо бережно хранить. Перед закладкой на хранение ее надо просушить в течение одного или нескольких часов. Рекомендуется ее хранить в пробирках, не заполненных доверху и закрытых ватой. Пробирки с пыльцой
помещают в эксикатор с хлористым кальцием. Эксикатор хранят при температуре 3-5°С. Перед использованием пыльцу следует проверить на всхожесть.
Опыление производится при помощи мягкой волосяной кисточки. При смене сорта пыльцы кисточку надо тоже менять или стерилизовать. При опылении цветков, изолированных в пакеты, применяют различные пыльцераспылители. Конец распылителя вводят в сделанный ножницами надрез пакета, впускают порцию пыльцы, а затем заклеивают отверстие лейкопластырем или изолентой. Для гарантии опыление повторяют через 1-2 дня.
После появления завязей изоляционные пакеты можно снять (через 10-14 дней). Но для сохранения плодов и семян необходимо бороться с энтомовредителями и другими повреждающими факторами. Для предотвращения разлета легких семян и смешивания с другими, а также повреждений птицами необходимо заранее закрыть завязи марлевыми мешочками.
Для различных видов древесных растений характерны разные технические приемы скрещиваний, часть из которых будет описана далее.
***
В заключение можно отметить, что гибридизация является весьма перспективным методом для получения соматического, адаптивного и генеративного гетерозиса. Применительно к лесным древесным породам этот метод особенно ценен для вегетативно размножаемых видов. С другой стороны, для перекрестно опыляемых видов этот метод требует доработки в плане последующего размножения полученных ценных генотипов. Если полученные хозяйственно ценные формы размножать обычным семенным путем, то существует большая вероятность потери отобранных ценных признаков.
Частично проблему могут решать биклоновые плантации, которые создаются из родительских пар с высокой СКС или введение в поликлоновые плантации растений с высокой ОКС. Однако и в этом случае требуется отбор среди потомков, так как имеется высокая вероятность получения гибридов не только с хорошими, но и нежелательными признаками.
Полное сохранение хозяйственно ценных признаков отобранных гибридов возможно только при вегетативном или апомиктическом размножении. Поэтому целесообразно было бы сосредоточить усилия на создании промышленных методов массового вегетативного (включая микроклональное) размножения хозяйственно ценных генотипов лесных древесных пород, полученных в результате направленной гибридизации. Кроме того, необходимо провести исследования по разработке методов искусственного получения апомиктов от хозяйственно ценных форм лесных древесных пород.

Вопросы для самопроверки
1. Что такое гибрид?
2. Перечислите цели гибридизации лесных древесных пород.
3. Какие задачи метод гибридизации позволяет решить при создании лесосеменных плантаций?
4. Охарактеризуйте конгруэнтные и инконгруэнтные скрещивания, их различия и значение.
5. Опишите характеристику комбинационных скрещиваний и принципы подбора пар при их использовании.
6. Опишите трансгрессивные скрещивания и их отличия от гетерозисных.
7. Охарактеризуйте явление гетерозиса, его типы и категории. Приведите гипотезы, объясняющие проявление гетерозиса.
8. Опишите методы гибридизации: простые и сложные.
9. Как получают инцухт-гетерозисные гибриды?
10. Укажите предпочтительные методы гибридизации для растений с разными системами размножения: самоопылителей, перекрестноопыляющихся и вегетативно размножающихся.
11. Приведите схемы диаллельных скрещиваний (полная, модифицированная, частичная, факториальная) и простое скрещивание пар; их отличия, достоинства и недостатки.
12. Какими особенностями должен обладать исходный материал для получения успешных скрещиваний?
13. Как технически может осуществляться гибридизация лесных древесных пород?
14. Перечислите достоинства и недостатки различных типов изоляторов для цветков древесных пород.
15. Охарактеризуйте режим хранения пыльцы и приемы искусственного опыления и получения семян у лесных древесных пород.

Интродукция является первичным способом оценки наследственной ценности организмов в новых условиях. Она нашла широкое применение в различных странах Старого и Нового Света. В России наибольшее число опытов по интродукции лесных древесных пород осуществлено в Европейской части страны. Ниже представлены некоторые подходы и результаты этой работы.
4.1. Теоретические предпосылки интродукции лесных древесных пород
Под интродукцией растений ботаники понимают целеустремленную деятельность человека по введению в культуру в данном естественно-историческом районе растений (родов, видов, подвидов, сортов и форм), ранее в нем не произраставших, а так же перенос их в культуру из местной породы. Однако для лесной интродукции более уместно определение В.И. Некрасова (1980), который рассматривает интродукцию как перенесение растений в новые природно-климатические условия за пределы естественного ареала или, при расширении площади искусственного выращивания растений, из района их испытания в культуре. Он не находит оснований считать перенос растений местной флоры в культуру интродукцией, т.к. он не связан с изменением географических условий, а приводит к изменениям агротехнических и биоценотических условий для роста растений. Действительно, если придерживаться ботанического понятия, то практически все лесные культуры, тем более связанные с изменением породного состава насаждения, следует считать интродукцией. Это заметно усложнит методологические аспекты интродукции лесных пород (И.И. Дроздов, 1998).
Тесно связаны с интродукцией понятия акклиматизации и натурализации растений. Акклиматизация — одна из форм интродукции растений, когда приспособление популяции к новым условиям обитания происходит за счет генетических изменений на основе местного естественного отбора индивидуумов, более чем исходные формы приспособленных к новым условиям обитания. Это характерно для случаев переноса растений в условия, значительно отличающиеся от естественного ареала. И поэтому, если интродукция влечет за собой огромные потери среди репродуцированной в новых условиях популяции и выживание лишь отдельных уклоняющихся генотипов из интродуцированного экотипа, говорят об акклиматизации. Натурализация подразумевает такой исход интродукции, когда новые формы растений легко произрастают и успешно репродуцируют в новых условиях, не изменяя своей генетической конституции. Как правило, это характерно для того случая, когда климатические и другие условия вполне благоприятны, отвечают биологическим особенностям интродуцента и он приживается без изменения генотипа.
При выборе исходного материала для интродукции приходится считаться с климатическими условиями произрастания. Знания климата района, откуда привлекается исходный материал, имеет существенное значение. Поэтому в интродукции древесных пород некоторое время господствовала теория климатических аналогов Г. Майра, которая связывала успех интродукции с непременным сходством климатических условий естественного и искусственного ареалов. Однако Н.И. Вавилов (1987) показал, что вопрос о климатических аналогах не может решаться в упрощенном виде. В целом он находит, что теория Г. Майра заслуживает большого внимания, поскольку дала немалые практические результаты. Однако в отношении древесных видов современная теория интродукции требует учета экотипов, сложности вида, на что Г. Майр не обратил внимание. По мнению Н.И. Вавилова (1987), климатические аналоги должны быть заменены экологическими аналогами, поскольку климатология и экология не равнозначны. Действительно, дальнейшее развитие теории интродукции подтвердили целесообразность эко-лого-исторического и эколого-географических методов. Прогнозирование эффекта интродукции признано немыслимым без экологического анализа.
Однако наиболее достоверные представления о возможностях интродукции дает опыт. Анализ предшествующего опыта интродукции — единственный способ определения степени адаптивности вида к новым условиям. В связи с этим широкое развитие получили методологические исследования по привлечению исходного материала. При подведении итогов интродукции, объяснении ее положительного или отрицательного опыта ориентируются на следующие методологические направления: сравнительный анализ климатических и эколого-географических условий естественного произрастания растений, привлекаемых в интродукцию, и новых мест их испытания; исторический и флорогенетический анализ растительности, из которой черпается материал для интродукции; изучение эколого-физиологических особенностей растений интродуцентов и изменчивости их анатомо-морфологического строения. Весьма важны результаты изучения сезонно-ритмических изменений в развитии вегетативных и генеративных структур для познания процесса приспособления интродуцентов.
Введение в лесные культуры экзотов, как правило, связано в дальнейшем с формированием интродукционной популяции. Вопросам создания интродукционных популяций на микро-эволюционном уровне уделил большое внимание В.И. Некрасов (1980). Использование для интродукции растений не случайных популяций, а обладающих формовым разнообразием, повышает гарантию успеха семенного размножения и получения более устойчивого потомства. Если интродуктор ставит своей задачей предсказать генетическую реакцию интродуцированных растений, то необходимо прежде всего изучить строение и состав исходных популяций и составляющих их
особей.
А.Я. Любавская (1982) отмечает, что успех интродукции данного вида часто определяется генетическим разнообразием вида. Интродукция экзотов — один из источников обогащения местной дендрофлоры и исходного материала для селекции. При интродукции ведется искусственный отбор по тем признакам, ради которых растения интродуцированы, а также по их устойчивости в новых условиях среды. Поэтому испытание интродуцентов должно быть поставлено на селекционно-генетическую основу. Для этого необходимо, чтобы в долгосрочной программе развития лесного хозяйства работа по введению интродуцентов была организованна на прочных селекционных принципах с организацией сортового семеноводства из отобранных по хозяйственно ценным признакам популяций и особей. Селекционно-генетическая оценка интродуцированных видов включает испытание межпопуляционной и внутрипопуляционной изменчивости экзотов и возможности отбора лучших популяций и форм для использования их при разведении в новых районах.
Придавая большое значение селекции и интродукции, В.И. Некрасов (1986) считает необходимым отдельно выделять маточные особи, которые при первичной интродукции и в последующих поколениях отличаются лучшими показателями роста, устойчивости И генеративного развития. От правильного выбора семян зависит успех интродукции. Поэтому разработка теоретических основ семеноводства интродуцированных древесных растений — насущная задача семеноводов. Семеноводство интродуцентов может развиваться успешно только на селекционной основе с использованием и постоянным развитием опыта географических культур. Райт Дж. (1978) отмечает, что интродукцию прежде осуществляли преимущественно семенами неизвестного, часто не лучшего происхождения. Установлено, что если интродуцированное дерево или насаждение дают 75 % продукции по сравнению с местной популяцией, то имеются основания продолжать проверку данного вида сравнительной оценкой различных климатипов и экотипов. А.Я. Любавская (1982) придает важную роль географическим культурам в практической интродукции. Изучение интродуцентов должно предусматривать накопление данных о влиянии географического происхождения и экологических условий на характер их роста и развития.
4.2. Особенности интродукции лесных древесных пород
Ботанические сады накопили ценные данные и значительный опыт по интродукционному испытанию многих древесных растений в коллекциях. Однако эти результаты можно считать предварительными для лесной интродукции, существенно отличающейся от интродукции видов в дендрариях, т.е. в изоляции, в отрыве от лесной среды. Объектом лесных интродукционных исследований является биогеоценоз или экосистема, главным компонентом которой служит насаждение. Здесь устойчивость интродуцируемого вида зависит всецело от его конкурентоспособности в лесном сообществе. Конечная цель лесной интродукции заключается в создании лесных ценозов, обеспечивающих наивысшую продуктивность древостоя главной породы.
Лесная интродукция (введение новых лесных пород в насаждения) обязательно связана с закладкой и выращиванием лесных культур. Экологической основой лесокультурного дела является лесная типология. Используя ее основные принципы, можно достаточно полно и комплексно представить характеристику лесорастительных условий на лесокультурной площади, выбрать тип культур, вид, метод и способ выращивания. Современное лесокультурное производство в решении вопросов лесовосстановления должно опираться на положения динамической типологии леса И.С. Мелехова (1984). Она охватывает практически весь цикл лесовосстановления со всеми возможными вариантами, которые позволяют выбрать и наиболее эффективно разместить лесные культуры в этом сложном природном процессе. Место их, в первую очередь, там, где процесс лесовосстановления затруднен или протекает медленно (через смену пород) (И.И.Дроздов, 1985, 1998).
Ценным индикатором при выборе типа лесных культур является коренной тип леса. Однако из этого не следует, что при создании лесных культур следует копировать природные типы леса. Вполне приемлемо здесь разумное использование других пород, в том числе интродуцированных, введение которых оправдано экологической и хозяйственной целесообразностью. В этом случае искусственные насаждения являются практически новыми компонентами динамической типологии леса. Формируется новый тип леса, который не вписывается в обычную типологию. Так, среди традиционных сосняков, ельников могут формироваться лиственичники, кедровники и другие как конечный итог лесной интродукции (И.И. Дроздов, 1985, 1998). Биогеоценотические сукцессии в ценозах, созданных из интродуцированнных пород, протекают значительно быстрее по сравнению с первичными, так как происходят на уже сформированных почвах. Вместе с тем механизм этих смен, будучи весьма сложными, к сожалению, еще слабо изучен (В.Б. Логгинов, 1988).
А.Я. Любавская (1982) считает, что трансформация насаждений изменением породного состава в ряде случаев практически необходима. При сплошных рубках естественное возобновление леса идет чаще всего через смену пород. В ряде случаев более выгодным становится выращивание в культурах экзотов. Интродукция растений перерастает в экономическую проблему. Испытание интродуцентов приобретает организованный характер и переводится на селекционно-генетическую основу.
Проектирование искусственных лесных ценозов из интродуцируемых пород трудно осуществить без детальных исследований особенностей формирования лесов из этих пород в естественных условиях. Поэтому экспедиционным путем с учетом накопленных материалов региональных исследований в натуре изучают типы леса наиболее важных лесообразующих пород, выявляют потенциальную продуктивность основных типов, определяют ведущие экологические и фитоценотические факторы, обеспечивающие потенциальный рост и развитие древостоев интродуцируемых растений в этих типах леса. На основе этого разрабатывают эколого-фитоценотические аналоги биогеоценоза — своеобразные природные эталоны будущих культурных лесонасаждений, обладающих продуктивностью, не уступающей потенциальной производительности естественных древостоев (П.И. Лапин и др., 1979).
Использование эколого-фитоценотических аналогов при интродукции древесных пород в лесохозяйственных целях конечно не означает простого копирования и переноса образцов природных древостоев в новые районы. Высокая продуктивность насаждений культурных ценозов, превышающая потенциальную производительность естественных, должна быть обеспечена подбором быстрорастущих, устойчивых экотипов и форм лесных древесных экзотов и соответствующим уровнем взаимодействия всех компонентов искусственного ценоза. При создании культур интродуцированных растений использование выявленных генетических особенностей обеспечивает более высокую степень их адаптации в ценозах, дальнейшее повышение производительности и качественного состава лесонасаждений.
С.Г. Спурри, Б.В. Барнес (1984) отмечают, что применяемый во всем мире при интродукции деревьев метод проб и ошибок отличается большой произвольностью, он дорог и требует много времени. Вместо этого надо использовать метод изучения генэкологической приспособляемости популяций разного происхождения. С этой целью лесоводы собирают семена желательных популяций, выращивают сеянцы и в молодом возрасте изучают различные потомства этих популяций. Эти эксперименты называют тестами на происхождение или источник семян, т.е. своеобразные географические посевы. Однако успех их во многом зависит от методики ранней диагностики генетических признаков.
Весьма важным моментом является изучение и анализ опыта интродукции деревьев и кустарников, накопленного ботаническими садами, научно-исследовательскими учреждениями, дендропарками, предприятиями лесного хозяйства и другими предприятиями. При этом первоочередное внимание уделяется изучению лесных культур древесных экзотов (П.И. Лапин и др., 1979). Учитывая важность этого аспекта лесной интродукции, на кафедре лесных культур Московского государственного университета леса на опыте 15-летних исследований разработаны методические рекомендации по изучению лесных культур интродуцированных пород (И.И. Дроздов, А.И. Янгутов, 1984).
Выявленные участки культур интродуцентов подлежат полному историческому, лесоводственному и лесокультурному описанию. При этом весьма важно установить происхождение исходного материала для культур: как они были созданы, как выращивали эти культуры. Далее культуры исследуют доступными для лесовода средствами. Дендрохроноклиматические исследования позволяют выявить, как росли экзоты в новых для них климатических условиях. Построенные при этом дендрошкалы позволяют получить объемную информацию при сравнительном анализе роста экзотов и местных пород (И.И. Дроздов, 1987). В пределах определенного климатического региона успешность выращивания той или иной древесной породы существенно зависит от почвенно-грунтовых условий. Изучение влияния этого фактора на рост новых пород позволяет определить критерии оценки почвенных условий и построить математическую модель роста интродуцентов (А.И. Янгутов, И.И. Дроздов, 1989).
Очень важно обратить внимание на устойчивость экзотов в изучаемом биогеоценозе не только в климатическом аспекте, но и по таким показателям, как наличие и характер заболеваний, повреждений вредителями, взаимоотношение с окружающей растительностью (древесной, травяной, грибами — микоризообразователями и др.), животным миром. Наблюдения за наличием и характером плодоношения экзотов в культурах позволяют судить о степени их акклиматизации и возможности использования насаждения для сбора семян местной репродукции. При исследовании возможности роста экзотов важно в первую очередь изучить динамику их сезонного развития и ритмику роста деревьев. В результате фенологических наблюдений устанавливают время наступления и продолжительность фенофаз (набухание почек, распускание их, рост побегов, развертывание хвоинок, летняя вегетация, одревеснение побегов и др.). Материалы эти, кроме научного, имеют для лесоводов практический интерес для определения оптимальных сроков посадки, уходов, подкормок и других агротехнических мероприятий.
Исследование особенностей индивидуального роста интродуцируемой породы в культурах ставит цель получить достаточно объективное представление о том, какими возможностями располагают эти деревья в онтогенезе. Степень изменчивости их основных таксационных показателей, закономерности роста дерева во времени весьма интересны для сравнения с такими же показателями породы В естественном ареале и особенностями роста местных лесных пород. Лесоводу они могут служить основанием для предварительного выбора типа лесных культур (компоненты насаждения, размещение, смешение, густота), метода и способа их выращивания, определения срока перевода культур в лесопокрытую площадь. Они позволяют провести отбор плюсовых деревьев в насаждении интродуцента. Большой интерес при искусственном лесовыращивании представляет динамика нарастания крон деревьев в фазах смыкания и жердняка. Сведения об этом позволяют прогнозировать сроки смыкания культур, правильно предусматривать их размещение и густоту.
Интродуктор, как правило, располагает сравнительно небольшим набором участков культур интродуцентов. Поэтому их следует изучить особенно тщательно и детально, чтобы не ошибиться в выборе экзота для последующего внедрения и свести к минимуму ошибки при проектировании будущих искусственных насаждений.
Рассматриваемые методы исследования позволяют получить качественные и количественные характеристики искусственных насаждений интродуцентов в различных условиях местопроизрастания, их взаимоотношения с местными лесными породами. Однако они не позволяют в полной мере раскрыть особенности формирования новых типов леса. Здесь необходимы специальные лесоводческие исследования, характер и направление которых определены положениями динамической типологии И.С. Мелехова (1999). Необходима специальная методика системного анализа состояния лесных экосистем, формируемых при сильном антропогенном воздействии (замена породы, регулирование состава и др.). Такие экспериментальные исследования позволяют в большей мере использовать теоретические аспекты динамической типологии леса, реализовывать их в качественных показателях, моделях выявленных взаимосвязей в искусственно формируемых насаждениях (И.И. Дроздов, 1985, 1998).
Долгосрочной программой по интродукции древесных пород в бывшем СССР предусматривалось проведение периодических (через 5-10 лет) единовременных учетов интродуцентов с детальным исследованием наиболее ценных объектов для уточнения ассортимента и лесоинтродукционного районирования по специальной разработанной ЦНИИЛГИС «Методике проведения единовременной инвентаризации интродуцированных лесных пород» (1986). Ее основные задачи:
1) выявление ассортимента древесных пород, интродуцированных в различные регионы европейской части СССР, а также площадей их посадок;
2) установление оптимальных географических и экологических условий, в которых соответствующие интродуценты дают наивысший хозяйственный эффект;
3) изучение состояния, продуктивности, агротехники культивирования, возраста и других параметров насаждений (состав, полнота), а также влияния биотических и абиотических факторов;
4) предварительная оценка возможности использования интродуцированных объектов как источников семенного и вегетативного материала;
5) разработка рекомендаций по обеспечению сохранности и использованию обследованных насаждений интродуцированных пород;
6) определение оптимальных типов лесных культур интродуцентов и технологии их выращивания.
При внедрении интродуцентов предусматривается этапность действий, связанная со степенью изученности показателей устойчивости и продуктивности тех или иных видов в отдельных регионах и наличием семенной базы. Вместе с тем строгое соблюдение этапности не всегда возможно. Ценные хорошо изученные виды-интродуценты при возможности сбора с них семян могут немедленно широко вводиться в производственные культуры. Одновременно с этими видами должна проводиться селекционная работа: отбор плюсовых деревьев и насаждений, создание клоновых архивов и испытательных культур, гибридизация, закладка семенных плантаций первого порядка, а после испытания потомства — плантаций второго порядка. Недостаточно испытанные в лесных культурах виды-интродуценты подлежат опытно-производственной проверке в культурах. Это, главным образом, экзоты, зарекомендовавшие себя в ботанических садах и перспективные по прогнозу для лесных культур. При недостатке семян закладке опытных культур может предшествовать создание соответствующей семенной базы.
Виды-интродуценты, представленные в ботанических садах единичными деревьями или небольшими биогруппами, а в лесных культурах единичными, случайными или неизвестного происхождения образцами, подлежат широкому первичному испытанию. Испытание таких видов или отдельных их происхождений проводится, главным образом, путем получения семян из естественного ареала. В определенных случаях оно возможно созданием специальных синтетических семенных плантаций.
4.3. Некоторые аспекты размножения
и внедрения интродуцентов
Успех интродукционной работы нередко зависит от способа размножения и выращивания растений. Поэтому лесоводу-интродуктору необходимо тщательно изучить биологию интродуцента, с тем чтобы определить правильную агротехнику искусственного выращивания его в новых условиях. Здесь перед лесоводом встают вопросы, связанные с подготовкой семян к посеву, сроками и способами их посева, защитой посевов от вредителей и болезней, с видами посадочного материала, агротехникой и сроками выращивания их, с типами лесных культур и агротехникой их выращивания, с лесоводственными приемами формирования насаждений и др. Необходимо определить место и значение в разведении экзота способов семенного и вегетативного размножения. Так, увлечение в 60-е годы прививкой сосны сибирской кедровой на сосну не оправдало себя. Сейчас пошли по пути разработки способов черенкования этой породы или сочетания прививок с последующем укоренением привоя (Р.Н. Матвеева, 1985; Г.Ш. Камалтинов, 1982).
Таким образом, при массовом введении в лесные культуры интродуцируемых пород необходимо решать ряд задач селекционно-генетического, растениеводческого, эколого-лесоводственного и экономического характера.
При организации интродукции Н.И. Вавилов исключительное значение придавал вопросам карантина, считая, что отсутствие карантинной службы может свести на нет самые успешные результаты экспедиционных работ как внутри страны, так и за рубежом. В связи с этим в составе ВНПО «Союзлесселекция», головной организацией которого являлся ЦНИИЛГиС, в 80-е годы была сделана попытка создания сети лесных карантинных питомников. Однако последующие годы реформ свели на нет это начинание.
***
В целом, результаты современных теоретических и методических разработок в области лесной интродукции свидетельствуют о ее реальных возможностях в решении актуальных задач лесного хозяйства. Надо, чтобы интродукция лесных пород была постоянно в поле зрения не одного поколения ученых и практиков без наблюдаемых ранее спадов, чреватых потерей сведений о происхождении культур тех или иных интродуцентов, разрывом в преемственности поколений, и в конечном счете потерей продуктивности лесов.
Обоснование экономической эффективности лесной интродукции можно считать решенным для пород быстрорастущих, которые в культурах промышленного назначения отличаются повышенной продуктивностью древесины при соответствующем ее качестве. Для интродуцентов, относительно медленно растущих, но имеющих немало преимуществ прижизненного функционирования (рекреационных, защитных, орехоплодных и др.), современная методика лесных экономических оценок фактически не работает. Это вызвано отсутствием количественных критериев экономической оценки целого ряда недревесных полезностей леса. И в этом направлении также требуется дальнейшая работа.
Вопросы для самопроверки
1. Что подразумевают под интродукцией и сопутствующими ей акклиматизацией и натурализацией растений?
2. Каковы отличительные особенности лесной интродукции?
3. Каковы принципиальные теоретические предпосылки плановой интродукции?
4. Охарактеризуйте методические аспекты изучения опыта интродукции.
5. Каковы фитоценотические особенности лесной интродукции?

Лесная селекция базируется на наследственной гетерогенности популяций древесных растений, которая позволяет вести отбор ценных генотипов. В последние годы в целях расширения селекционной базы наряду с традиционными способами повышения генетического разнообразия, такими как гибридизация и интродукция, успешно внедряются новые экспериментальные методы, основанные на достижениях генетики, эмбриологии и цитологии растений. Среди таких методов ведущее место занимает индуцированный мутагенез, полиплоидия и культура клеточных тканей и клеток.
5.1. Общие положения по использованию
мутагенеза, полиплоидии и культуры тканей
в селекции лесных древесных пород
Мутации, как известно, являются одним из основных факторов эволюции живой природы. Спонтанные мутации, возникающие как следствие естественных мутагенов, в гомозиготном состоянии проявляются фенотипически и немедленно подвергаются естественному отбору, а в гетерозиготном состоянии могут долго сохраняться и накапливаться в популяции.
Мутации, возникающие в половых клетках, называются генеративными, и передаются новому поколению. Мутации, происходящие в любых других клетках организма, называются соматическими; они наследуются только при вегетативном размножении. Один из видов соматической мутации у древесных растений почковые мутации, возникающие, в меристемных клетках точки роста стебля (предполагают, что благодаря этой мутации возникли пирамидальные тополя), Различают генные, хромосомные и геномные мутации.
Генные мутации — изменения, возникающие вследствие перестройки в молекуле ДНК чаще всего при замене одного или нескольких нуклеотидов в пределах гена. Это приводит к изменению строения молекул белков и появлению новых признаков и свойств (например, гроздешишечные формы у хвойных, «ведьмины метлы», альбиносы, махровость у цветков, образование летальных генов и т.д.).
Хромосомные мутации связаны со структурными изменениями хромосом, при которых меняется положение участков хромосом, нарушаются существующие и возникают новые группы сцепления.
Геномные мутации — это изменение числа хромосом. Они могут быть вызваны нерасхождением хромосом при делении. Особенно важными для селекции являются геномные мутации, возникающие из-за нерасхождения хромосом при мейозе, так как следствием может быть появление нового набора хромосом у гамет. Геномные мутации могут быть представлены гаплоидами (с хромосомной мутацией), анеуплоидами (с лишней или недостающей хромосомой), полиплоидами (с кратным увеличением хромосом).
Спонтанные мутации возникают редко, носят случайный, ненаправленный характера Они приводят к появлению „необычных форм древесных растений. В природе у древесных растений кроме мутаций, приводящих к изменению формы кроны,(встречаются хлорофилльные мутации, мутации с морфологическими нарушениями листьев (рассеченные листья) и т.п. Такое мутантное дерево обычно произрастает в парках или садах, так как в лесу вероятность выживания его очень низкая. Причиной появления около 50% спонтанных мутаций у древесных пород считается естественная радиоактивность. Значительно больший интерес с точки зрения получения практических результатов имеет направленное получение мутаций, которое базируется на раскрытии молекулярных механизмов мутаций, управлении этими мутациями и получении форм живых организмов с заранее заданными признаками и свойствами. В селекции растений — это чаще всего выведение полиплоидных форм.
Полиплоидия, или кратное увеличение гаплоидного набора хромосом, вызванное геномной мутацией, — обычное явление в биологии. По существу, в жизненном цикле высших растений спорофит является полиплоидом по отношению к гаметофиту, так как клетки его содержат удвоенный (диплоидный) набор хромосом.
У древесных растений более трети всех родов имеют полиплоидные виды. У голосеменных древесных растений естественная полиплоидия наблюдается очень редко. Истинным полиплоидом среди древесных форм голосеменных является секвойя вечнозеленая {Sequoia sempervirens), у которой клетки спорофита содержат 66 хромосом.
Иная картина наблюдается у лиственных пород. По мнению некоторых исследователей, виды
Дата добавления: 2017-01-13; просмотров: 3235;