Индивидуальный отбор у перекрестноопыляющихся растений
Вследствие постоянного переопыления и расщепления признаков сорта у перекрестноопыляющихся растений нельзя вывести путем однократного индивидуального отбора. Поэтому в селекции таких растений применяют многократный или непрерывный (в каждом поколении) индивидуальный отбор. Эти виды отбора являются улучшающими и они широко используются в сельском хозяйстве. При этом имеют дело не с отдельными растениями, а с семьями. Семья — это потомство одного отобранного у перекрестноопылителей растения. Урожай семян растений одной семьи также высевают раздельно по семьям. Семьи сравнивают между собой, худшие бракуют, из лучших снова отбирают лучшие растения для последующего размножения и т. д. Таким образом, сущность данного метода заключается в повторяемом отборе лучших растений из лучших семей.
В отличие от самоопыляющихся и вегетативно размножаемых растений потомство перекрестноопыляющихся растений формируется в результате переопыления, то есть на основе объединения материнской и отцовской наследственности. Поэтому в селекционной работе с перекрестноопыляющимися растениями важно не только правильно вести отбор лучших материнских растений, но и подбирать такие отцовские растения — опылители, которые не ухудшали бы, а по возможности улучшали наследственные качества будущего потомства. В общем виде задача сводится к необходимости контролировать отбор не только по материнской, но и по отцовской линии. Исходя из этого, в практике селекции сельскохозяйственных растений, а в последние десятилетия и лесных древесных пород, используют два основных варианта многократного индивидуального отбора: индивидуально-семейный и семейно-групповой.
Индивидуально-семейный отбор заключается в том, что после отбора лучших растений в популяции исходного материала потомство каждого лучшего растения, называемого семьей, размещают изолированно от остальных потомств. Этот метод первоначально был применен при отборе перекрестноопыляющихся сельскохозяйственных культур. Его сущность описана Г.В. Гуляевым и др.(1987) и Ю.Л. Гужовым и др.(1991). Схема данного отбора представлена на рис. 2.9. Здесь при изолированном размещении потомств в лучших семьях повторно отбирают лучшие растения. По сравнению с первым отбором в исходном материале второй отбор имеет то преимущество, что отобранные лучшие растения опыляются пыльцой не всех испытуемых растений данной популяции, а только в пределах данной семьи, которая в целом показала хорошие качества. Контролируемое переопыление лучших растений приводит к некоторой гомозиготизации материала, которая, с одной стороны, позволяет более эффективно вести отбор по отношению к аддитивным генным эффектам, но с другой — вызывает частичную инцухт-депрессию, что следует учесть при дальнейшем испытании потомств.
Потомства лучших растений второго индивидуального отбора снова оценивают при изолированном их размещении по семьям, а отбор ведут по потомствам. Лучшие потомства в пределах каждой семьи объединяют, испытывают и размножают.
Метод индивидуально-семейного отбора сочетает преимущества индивидуального и массового отборов. Двукратным индивидуальным отбором можно найти наиболее ценные генотипы, а возможные отрицательные последствия, возникающие в результате сравнительно близкородственного размножения при индивидуальном отборе, снимаются путем объединения лучших потомств каждой семьи как при массовом отборе.
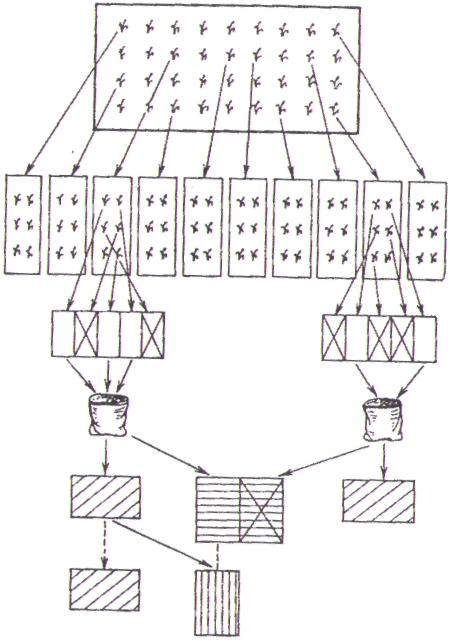
|
| Исходный материал. Отбор лучших растений |
| 1 -й год |
| Посев по семьям на изолированных делянках. Отбор лучших растений в семьях Оценка семей. Объединение лучших семей |
| 2-й год |
| 3-й год |
| Размножение и испытание лучших семей |
| 4-6-й годы |
| 7—10-й годы |
| Государственное сортоиспытание |
Рис.2.9. Схема индивидуально-семейного отбора
Индивидуально-семейный отбор применяется и для лесных древесных пород (Э. Ромедер, Г. Шенбах, 1962; И. Добринов, 1983). В качестве примера можно рассмотреть, как он проводится в Германии. В коренном исходном насаждении отбирают 60-100 плюсовых деревьев. В семенной год собирают семена или плоды с каждого дерева и высевают раздельно следующей весной. Как только растения в питомнике достигнут определенного размера, их пересаживают на опытные делянки достаточных размеров для выращивания растений в течение всего срока испытания. Повторность делянок минимум трехкратная. Срок испытания у разных пород может быть разным: у ели 20-30 лет, у березы и ольхи 15-20 лет и т. д. Исследуется рост, феноразвитие, устойчивость, качество стволов и древесины и другие хозяйственно ценные признаки. В конце периода испытания из каждых 100 потомств
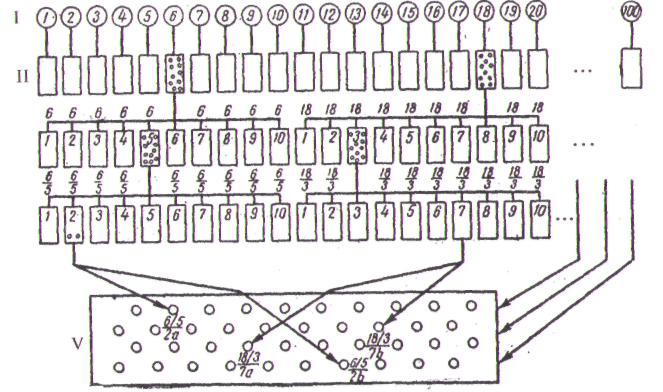
Рис. 2.10.Схема селекции семейств или отборных деревьев:
I — материнские деревья; II — потомства отдельных деревьев; III — испытание А
(по 10 лучших деревьев из II); IV - испытание Б (по 10 лучших деревьев из А);
V - семенная плантация (по 2 лучших дерева — а и b — из 10 лучших семей Б).
(Э.Ромедер, Г. Шёнбах, 1962.)
выбирают только 10 наиболее соответствующих селекционным целям. Из каждого отобранного потомства отбирают по 10 наиболее продуктивных деревьев и заготавливают с них семена или плоды. Схема для этого вида отбора приведена на рис. 2.10.
Семена каждого дерева высевают на одной из 100 опытных делянок с соответствующими повторностями. Это называется испытанием А. Оно продолжается 15-30 лет. В конце данного периода вновь устанавливают 10 лучших потомств и от каждого из них выбирают по 10 лучших деревьев. Собранные семена вновь высевают на 100 опытных делянках с повторностями. Это испытание Б. Через 15-20 лет выявляют 10 лучших семейств и от каждого из них выбирают по два лучших дерева. От этих деревьев (всего 20) берут черенки для прививки. Привитые растения высаживают на плантацию таким образом, чтобы близкородственные особи не росли рядом. При таком способе отбора идет сознательное сужение наследственных основ и может наблюдаться депрессия роста, вызываемая инцухтом. Если это будет замечено (для чего в испытании необходимо в качестве контроля использовать торговый семенной материал), испытания А и Б следует прекратить и довольствоваться достигнутым результатом.
Описанный способ очень длительный, однако Мюнхенский институт семеноведения и селекции лесных культур в 1935 г. приступил к селекции ели этим способом. Первые результаты оказались, по данным Э. Ромедера и Г. Шенбаха (1962), обнадеживающими. В последующих сводках по селекции лесных древесных пород в Германии (Н. Hattemer u. а., 1993) результаты опыта не упоминаются. Недостаток этой схемы, по-видимому, заключается и в том, что в конечном итоге рекомендуется на лесосеменной плантации использовать только 20 растений. Известно, что в последние годы это количество уже не считается достаточным (см. гл. 8).
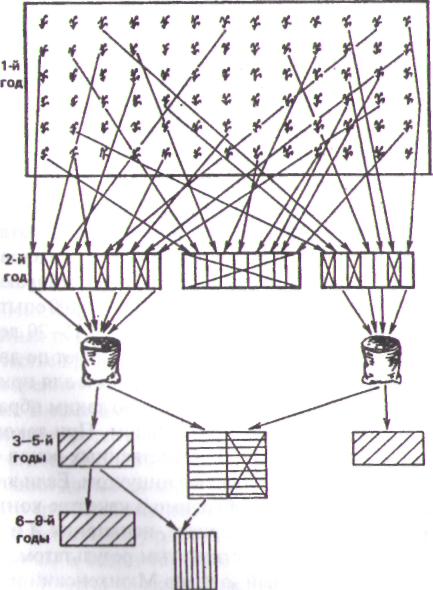
|
| Исходный материал. Отбор лучших растений и распределение их по сходным группам |
| Посев групп семей на изолированных участках. Объединение лучших семей |
| Размножение и испытание лучших групп семей |
| Размножение лучших групп семей. Государственно* сортоиспытание |
| Рис. 2.11. Схема семейно-группового отбора |
Семейно-групповой отбор производят по схеме на рис. 2.11. Ее описание приведено в работах Г.В. Гуляева и др. (1987) и Ю. Гужова и др. (1991). В популяции исходного материала отбирают лучшие растения. В соответствии с определенными физиологическими или морфологическими признаками (рост, прямизна ствола, характер ветвления, устойчивость к болезням и т. п.) их объединяют, соблюдая при этом принцип индивидуального отбора. Создают несколько групп (3-4 и более). Потомства лучших растений каждой группы семей размещают на изолированном участке. Внутри группы отбирают лучшие потомства, а остальные выбраковывают. Эти потомства переопыляются между собой в пределах данной группы.
Таким образом, вероятность близкородственного размножения уменьшается, а переопыление происходит в рамках сходных семей. Лучшие потомства каждой группы объединяют по принципу массового отбора. Смеси из лучших потомств каждой группы испытывают и размножают. В ряде случаев в пределах группы отбор проводят дважды.
При семейно-групповом отборе сохраняются преимущества направленного переопыления, но снижается опасность близкородственного размножения и связанной с ним инцухт-депрессии. В то же время выравненность селекционного материала достигается медленнее, чем при индивидуально-семейном отборе.
Этот вид отбора может применяться и к лесным древесным породам (И. Добринов,1983; L. Paule, 1989). Однако, как и индивидуально-семейный отбор, этот вид отбора является очень продолжительным и для основных лесных пород требует 70-100 лет и более.
***
Можно отметить, что до последнего времени приоритетными направлениями в селекции лесных древесных пород был массовый отбор климатипов. Это направление, очевидно, будет актуальным еще достаточно длительный период времени. Обусловлено это тем, что по мере применения существующих рекомендаций выясняются вопросы, которые требуют уточнения и детализации. Особенного внимания требует вопрос о значении экологического происхождения семян. Считается, и на наш взгляд вполне справедливо, что при лесокультурных мероприятиях необходимо учитывать этот параметр. Однако еще не хватает экспериментальных данных, которые с достаточной полнотой оценивали бы потери, которые может понести хозяйство при игнорировании экологического происхождения семян. Все еще дискуссионным остается вопрос об использовании массового и индивидуального отборов в пределах лесосеменных районов. Отбор плюсовых насаждений и плюсовых деревьев является только первым этапом сложной системы селекционного семеноводства. Необходимы длительные и трудоемкие работы, связанные с испытанием их потомств, чтобы достичь определенного эффекта.
Иногда встречается путаница в терминологии. Некоторые полагают, что отбор плюсовых деревьев и есть индивидуальный отбор, что, конечно, не совсем так. Все зависит от дальнейших схем селекции, а это могут быть и схемы индивидуального отбора (индивидуально семейный и семейно-групповой), и схемы массового отбора (общий сбор семян с плюсовых деревьев без дальнейшего учета, с какого дерева собраны эти семена).
Таким образом, несмотря на то, что отбор как метод используется очень давно, все еще остаются проблемы, которые требуют своего решения. Следует отметить также, что поставленные вопросы касались главным образом основных лесообразующих пород, однако те же проблемы стоят и перед интродуцентами и перед другими хозяйственно важными древесными породами.

Вопросы для самопроверки
1. Охарактеризуйте основные виды естественного отбора.
2. Назовите виды искусственного отбора и дайте их определения.
3. Раскройте достоинства и недостатки массового отбора. Опишите опыты Иоганнсена по отбору в гомозиготных популяциях.
4. Дайте определение позитивного, негативного и отрицательного отбора. Какие методы позитивного отбора применяются в лесном хозяйстве.
5. Покажите значение географического происхождения семян для роста и развития растений, а также проявление характеристик климатипов по О. Г. Капперу.
6. Какие основные принципы лесорастительного районирования были приняты в 1982 г.? Остались ли они актуальными в настоящее время?
7. Оцените значение эдафотипов для роста и развития лесных древесных пород.
8. Назовите группы экотипов дуба черешчатого и сосны обыкновенной, выделенные М. М. Вересиным. Каково их значение для лесоразведения?
9. В чем заключается значение отбора лучших (плюсовых) насаждений для лесоразведения?
10. В чем заключается значение отбора лучших (плюсовых) деревьев для лесоразведения?
11. В чем заключается значение сортировки растений по величине и семян по массе для лесовыращивания?
12. Дайте общую характеристику метода индивидуального отбора.
13. В чем состоит сущность метода педигри и клонового отбора?
14. Охарактеризуйте метод индивидуального отбора у перекрестно опыляющихся растений.
15. В чем заключается сущность индивидуально-семейного отбора? Приведите примеры и схему этого отбора, предложенную немецкими исследователями для лесных древесных пород.
16. Раскройте сущность и схему семейно-группового отбора.
17. Охарактеризуйте основные проблемы отбора как метода селекции.
Глава 3.
Методы селекции. Гибридизация
При гибридизации получают новые комбинации генов, которые не встречаются в исходном родительском материале. Генетики склонны считать, что исходные родители представляют полюса разнообразия каких-либо признаков, и гибриды только заполняют пропуски между этими полюсами (Wagner, 1969, цит. по В. Zobel, J. Talbert, 1984). Это положение вытекает из менделевской генетики (Э. Баур, 1922). Вагнер только еще раз подчеркивает, что ничего «нового» при гибридизации не создается и она только противодействует крайностям в характеристиках, которые вызываются эволюционными факторами, такими как мутация, отбор, генетический дрейф.
Однако, применяя метод половой гибридизации, селекционеры могут создавать и отбирать такие формы лесных древесных пород, которые по тем или иным причинам не встречаются в природе. На огромный потенциал гибридизации для лесных древесных пород указывали как русские ученые (С.З. Курдиани, Н.В. Сукачев, А.С. Яблоков, А.В. Альбенский и другие, упоминавшиеся в первой главе), так и зарубежные (Stockwell, Righter, 1947; Righter, 1955; Duffield, Snyder, 1958, цит. по В. Zobel, J. Talbert, 1984).
3.1. Некоторые общие положения гибридизации
Некоторые сведения о начальных этапах использования гибридизации растений, включая и лесные древесные породы, приводит Л. Пауле (табл. 3.1). Имена указанных в данной таблице исследователей широко известны в научном мире.
Необходимо кратко остановиться на самом понятии «гибрид». Первоначально гибридами или бастардами называли организмы, полученные в результате скрещивания двух видов. В современном понимании гибрид — это гетерозиготная особь, возникающая в результате скрещивания генетически различных родительских форм или генотипов, т.е. любой гетерозиготный организм, независимо от его происхождения (Р. Ригер, А. Михаэлис, 1967; Г.В. Гуляев,
Таблица 3.1
Дата добавления: 2017-01-13; просмотров: 1830;