Роль ретикулярной формации в угнетении условных рефлексов.
В последние годы это положение доказано электрофизиологическим исследованием. Установлено, что у кошки под влиянием электрического раздражения лапы, вызывающего оборонительную реакцию, кора претерпевает общее повышение активности, которое осциллографически выражается в устранении медленных волн и в усиленной продукции бета-волн — в так называемой десинхронизации. В этот момент в ответ на раздражения не возникают первичные потенциалы и не наблюдается последующего усложнения их быстрыми потенциалами. Ройтбак (8) показал, что при электрическом раздражении кожи как во время раздражения, так и некоторое время после него, измеряемое иногда десятками секунд, звуки не вызывают в слуховой области первичных потенциалов (рис. 170).
Эрнандец-Пеон и сотрудники (18) показали, что вызываемые зрительным раздражением потенциалы в коре, в боковом коленчатом теле и ретикулярной формации среднего мозга частично или целиком исчезают при звуковых и обонятельных раздражениях, вызвавших ориентировочную реакцию или, как говорят авторы, реакцию внимания.
Далее, установлено, что после прямого раздражения ретикулярной формации ствола мозга, особенно при ее возбуждении от сильного электрокожного раздражения, одновременно с торможением внешних реакций на другие раздражения, происходит депрессия афферентных импульсов по восходящим путям в течение многих десятков секунд по прекращении раздражения (Эрнандец-Пеон с сотрудниками, 18, 19). Это торможение происходит в афферентном пути между первым и вторым чувствительными нейронами — в ядрах спинного и продолговатого мозга (Хагбарт и Керр, 20, Ройтбак, 8). Гершуни, например, показал, что у кошки во время акта еды, при сильных механических раздражениях угнетается поток афферентных импульсов на различных уровнях слуховой системы (21).
На основании подобных наблюдений можно заключить, что угнетение условного рефлекса под влиянием необычных раздражений происходит главным образом не вследствие коркового торможения, а благодаря блокаде афферентных импульсов в ядрах ствола мозга. Предполагается, что в стволе мозга — в ретикулярной формации — существует механизм, регулирующий афферентную импульсацию (Эрнандец-Пеон с сотр., 18; Гершуни, 21). Эта регуляция должна осуществляться путем передачи импульсов возбуждения из клеток этого механизма через их аксоны к чувствительным ядрам. Здесь они, должно быть, активируют дендритную массу, вызывая медленные потенциалы, которые могут произвести анэлектротоническое угнетение синапсов на телах соответствующих нейронов и тем сгшым прекратить передачу возбуждения через эти ядра в кору больших пслу/гарии.
Активация ретикулярной формации первично происходит под влиянием афферентной импульсации из рецепторов. Однако ретикулярная формация может активироваться и вторично под влиянием центрифугальной корковой импульсации (см. подробно Нарикашвили, 22). Это хорошо проявляется при раздражении коры. Возникшая при этом импульсация по кортикофугальным путям влияет на ретикулярную формацию, производя через нее как блокаду, так и облегчение проведения афферентных импульсов (Эрнандец-Пеон и Хагбарт, 19). Следовательно, можно предположить, что при возбуждении коры афферентными импульсами по специфическим путям в ней могут воз-
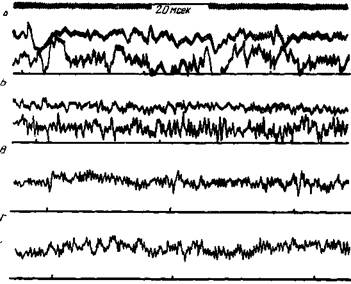
Рис. 170. Явление устранения первичных потенциалов в слуховом анализаторе при электрической раздражении кожи
Собака с отводящими электродами в слуховой (верхние кривые) и кожной (нижние кривые) областях коры. А — эффекты трех звуковых толчков при спокойном состоянии. Момент каждого толчка отмечается черточкой. Б — эффекты того же звукового раздражителя во время электрического раздражения кожи передней лапы, вызвавшего сильное движение. В — потенциалы отводятся только в слуховой области. Эффекты трех звуковых толчков. Г — аффекты того же звукового раздражения, когда собака ела мясо (Ройтбак)
будиться кортикофугалыше пути, которые в свою очередь могут влиять на ретикулярную формацию, вызывая торможение или облегчение проведения афферентных импульсов. На большое значение ретикулярной блокады проведения афферентных импульсов на уровне ствола в эффекте внешнего торможения указывают и поведенческие опыты с одновременной регистрацией ответных потенциалов. Если, например, нормальной кошке с электродами, выявленными в зрительной или слуховой системе, показать мышь или дать вдыхать запах рыбы, то в течение всего времени, пока кошка эмоционально возбуждена и настороженно следит за мышью или вдыхает запах рыбы, первичные ответы на звук или свет оказываются угнетенными (Эрнандец-Пеон, Шеррер и Жуве, 23). После устранения этих раздражений ответы на звук или свет вновь восстанавливаются до первоначальной амплитуды. Важно отметить, что угнетаются не только корковые ответы, но и ответы, возникающие на всех подкорковых уровнях афферентной системы. Характерно, если эти, в данном случае, пищевые раздражения, повторяются несколько раз или продолжаются длительное время без того, чтобы овладеть пищей, то происходит угасание натурального условного рефлекса захвата мыши или рыбы. Поэтому после этого амплитуда первичных ответов на звук или на свет восстанавливается до первоначальной величины. Угнетение первичных ответов той или другой афферентной системы получается при любом более или менее сильном добавочном периферическом раздражении, а также при непосредственном электрическом раздражении ретикулярной формации. Отсюда и делалось заключение о том, что наблюдаемое в поведенческих опытах угнетение ответов под влиянием экстрараздражений связано с активацией ретикулярной формации (Эрнандец-Пеон, 24).
В пользу ретикулярного происхождения внешнего торможения говорят и результаты опытов с повторным раздражением ретикулярной формации. Если несколько раз друг за другом повторить периферическое раздражение, которое активирует ретикулярную формацию, или повторно электрически
раздражать эту формацию (Гликман и Фельдман, 25), то угнетающее влияние последней значительно ослабевает и прекращается. Оно возникает вновь, если увеличить интервал между повторными раздражениями.
Таким образом, как в случае условных натуральных раздражителей, так и при более или менее сильном периферическом раздражении в вышеописанных поведенческих опытах, вызывается наиболее выраженная ориентировочная реакция и оказывает наибольшее угнетающее действие на первичные корковые ответы при первых применениях. Значит, в основе внешнего торможения лежит более или менее генерализованная активация ретикулярной формации, которая может наступать под влиянием как афферентных импульсов, так и кортикофугалъных возбуждений, в обоих случаях через коллатерали на уровне ствола головного мозга.
Дата добавления: 2017-01-13; просмотров: 818;