Лабиринтные рецепторы.
Ориентация в пространстве при закрытых глазах может протекать более или менее нормально посредством одних лабиринтных рецепторов, т.е. при участии одного вестибулярного анализатора, следовательно, без непосредственного участия не только анализаторов обонятельного, слухового, кожного, но и без прямого участия двигательного анализатора. Так, например, если животное — собаку или кошку — посадить с завязанными глазами в клетку и перенести клетку в другой угол комнаты на расстояние нескольких метров и там накормить, а затем отнести таким же путем обратно, то спустя несколько минут животное сдмо может повторить этот путь не только с открытыми глазами, но и с закрытыми, если только оно привыкло в достаточной степени к ношению маски (рис. 113). Этими опытами мы точно установили, что активное передвижение животного к месту пищи при закрытых глазах происходит исключительно посредством лабиринтных рецепторов. В данных опытах завязывание глаз исключало возможность зрительных восприятий. Так как во время переноса животное находилось в клетке без движения, то были исключены также кинестетические раздражения конечностей, обычно наступающие при шагательных движениях. Роль обонятельных раздражений устранялась или протиранием пола бензином, или перерезкой обонятельных нервов. Это доказано и следующего рода опытами: если перенести животное с завязанными глазами сначала в один угол и там подкормить, а затем в другой угол без подкармливания, то через несколько минут, если дать свободу, животное пойдет сразу к месту подкармливания, а к другому месту не пойдет. Бывает и так, что животное по пути может натолкнуться на какой-либо предмет и потерять ориентацию. Однако, если после этого открыть глаза, оно сразу побежит к месту пищи. Из этого видно, что от одного переноса в клетке к новому месту пищи при завязанных глазах животное приобретает образ пути переноса (Беритов, 5). Более того, если собаку с закрытыми глазами перенести сначала налево наЗ—4да в сторону кормушки 1 и затем, не накормив, отнести направо, как показано на рис. 114, и там подкормить, а после этого возвратить по тому же пути в клетку, то собака спустя некоторое время сама может пойти к месту пищи, не по следам пройденного пути, а прямо по короткому пути, по которому ее не водили. Таким образом, ясно, что животное с закрытыми глазами, на основании лабиринтной рецепции, точно локализует место пищи, куда его приводят или переносят в клетке (Беритов, 1, 5).
Посредством лабиринтной рецепции не только воспринимается пройденный путь к определенному месту пищи, но точно локализуется и конец пути — место пищи в экспериментальной комнате. Вследствие этого, если собаку отвести или перенести в клетке в любое место комнаты, она может оттуда
| Рис. 113. Ориентированное пищевое поведение после однократного переноса к месту пищи Собака Джильда Глаза завязаны Сплошные линии обозначают пути переноса животного в клетке, а штриховые линии — свободного передвижения. По пути а переносим собаку в клетке к ормушке 1 и подкармливаем Обычный подход к кормушке был загорожен. Через 2 мин она сама идет по пути а1 небольшими зигзагами, наталкивается на заграждение и поворачивает назад, б — путь переноса за кормушку 2, где ее подкармливают б1 — через минуту сама идет в том же направлении, наталкивается на ширму экспериментального стола и отсюда идет к кормушке 1, в — после снятия повязки идет прямо за кормушку 2 | 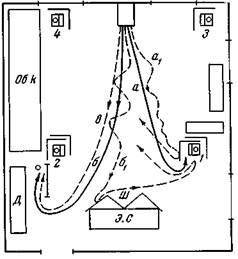
|
| Рис. 114. Ориентированное пищевое поведение по новому пути Собака Джильда с завязанными глазами, а — путь переноса в клетке по ломаной линии к месту, где ее подкармливают из миски, и обратно в клетку по тому же пути, б — через минуту открываем клетку и собака сама идет к миске по более короткому пути после еды собаку отводим обратно по пути а, в — через две мин выпускаем из клетки и она идет прямо вперед к экспериментальному столу, наталкивается здесь на ширму, прикрывающую экспериментальный стол, и поворачивает налево, отсюда отводим в клетку, а затем вновь по пути а, г — через минуту вновь выпускаем из клетки и теперь она идет прямо к миске | 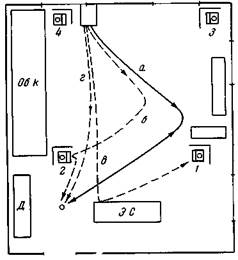
|
| Рис. 115. Ориентированное пищевое поведение собаки Дики с завязанными глазами Третий день работы в экспериментальной комнате После того, как со баку девять раз перенесли в клетке по пути АБ и подкормили, выводя из клетки, стали переносить ее в другие места, не давая еды Каждый раз она сама выходила из клетки и шла прямо к месту пищи Обозначения те же, что и на рис 114, а и a1 — свободное передвижение после выхода из клетки на месте обычного пребывания, б — после переноса по обычному пути А Б, в — после переноса по необычному пути АВ, г — после необычного переноса по пути AI , о — после переноса по пути А до поворота 0 | 
|
|
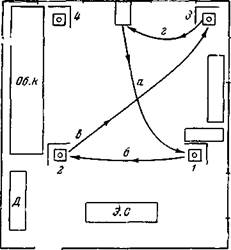
| Рис. 116. Схема пути пищевого поведения собак Джек и Акбар при подкармливании из трех кормушек (по опытам Т. Ониани) а, б, виг указывают на последовательность передвижения собаки с кормлением из кормушек 1, 2 и з |
прямо пойти к месту пищи, производя при этом правильно все необходимые повороты (рис. 115) (Беритов, 1, 5). В рассмотренных выше случаях собака с закрытыми глазами бежала по довольно простому и короткому пути. Но тот же результат был получен, когда путь к кормушке значительно усложнили. Так, например, при звонке собаку Джека отводили за ошейник сначала к кормушке 1, затем по окончании еды ее вели к кормушке 2, где также подкармливали, наконец отводили к кормушке 3 и после подкармливания из этой кормушки возвращали в клетку (рис. 116). После двух таких насильственных отведений и подкармливании собака сама побежала к кормушкам, когда ее вывели из клетки, и обошла их в том же порядке, как при насильственном отведении. Когда мы закрыли собаке глаза, надев на нее маску, она и тогда пошла к этим кормушкам, и притом в том же порядке, как и при открытых глазах. Собака была предварительно хорошо приучена к маске, и она проделала свой обычный путь к кормушкам уже при первом испытании звонка (Ониани, 6).
По характеру этих наблюдений нужно было предположить, что собака имеет зрительные образы местоположения кормушек в экспериментальной обстановке и только поэтому она в состоянии обходить данные кормушки с закрытыми глазами. Дальнейшие опыты показали, что это предположение не совсем правильно. Другую собаку (Акбар), предварительно приученную к ношению маски, мы стали отводить в маске к кормушкам в том же порядке, как и предыдущую собаку. Уже после двух насильственных отведений при звонке собака начинала выходить из клетки на звонок. В первый опытный день при звонке она не была в состоянии обойти все кормушки в обычном порядке. Кроме того, она путала порядок. Но уже на другой день в самом начале опытов, как только ее усадили в клетку (надев маску), она сама вышла из нее и пошла к кормушкам, обошла их в том же порядке, в каком ее отводили при подкармливании. Кормушка не открывалась и поэтому, постояв немного, собака шла дальше к следующей кормушке, причем она шла коротким путем спокойно и после посещения кормушки 3 сама возвратилась в клетку (Ониани, 7).
Только на третий день после двадцатикратного отведения к кормушкам собака начала и при звонке обходить кормушки в обычном порядке. Она шла сначала прямо с поднятой головой к кормушке 1 и, подойдя близко, опускала голову и производила искательные движения с обнюхиванием пола. При этом она высоко поднимала передние лапы, как обычно делала при еде из кормушки, так как во время еды она передними лапами опирается на ступень кормушки, которая возвышается на 10 см над полом. Затем подходила к кормушкам 2 и 3, производя искательные движения и поднимание лапы (Ониани, 7).
Изэтих наблюдений видно, что собака может производить ориентированные движения с закрытыми глазами не только к одной кормушке, но и к ряду их, и притом не только на основе зрительных образов местонахождения кормушек, но и путем создания этих образов посредством лабиринтных рецепторов.
Понятно, только на первой стадии образования пищевого поведения последнее регулируется, как уже говорилось, возникшим при этом зрительным или вестибулярным образом местонахождения кормушек и проецированием их в определенном месте экспериментальной комнаты. По-видимому, эти образы воспроизводятся и проецируются всякий раз при пищевых сигналах, а также и при восприятии других предметов экспериментальной комнаты. Благодаря проецированию образа местонахождения кормушек собака может пойти к ним из любого участка этой комнаты.
Но после многократного кормления из ряда кормушек в определенном порядке при каком-либо звуковом сигнале вырабатывается автоматизированное условнорефлекторное хождение на этот сигнал к данному ряду кормушек.
Возникший в зрячем состоянии образ местонахождения кормушки и его регулирующее значение в пищевом поведении животного сохраняется и после закрытия глаз. Более того, этот образ местонахождения кормушки создается и без участия зрения — при завязанных глазах, значит, на основе лабиринтной рецепции.
Отсюда следует, что у собак, как и у кошек, пространственная локализация или пространственное проецирование объектов, а также ориентированное движение к этим объектам не является исключительной функцией зрительного анализатора. Каждый раз при передвижениях собаки из клетки к кормушке и вообще от предмета к предмету, даже при наличии зрения, у нее в пространственном проецировании воспринятых предметов принимает участие и вестибулярный анализатор. Этим, понятно, объясняется, что пространственная ориентация в окружающей обстановке, приобретенная при участии вестибулярного аппарата, сохраняется почти полностью и после выключения зрения.
Итак, раздражения лабиринтных рецепторов при передвижениях животного являются существенным фактором пространственной ориентации в окружающей среде. Из этого следует, что слепое или нормальное животное с завязанными глазами (или при абсолютной темноте) может хорошо ориентироваться только при помощи лабиринтных ощущений.
Безлабиринтное животное (с двусторонним разрушением лабиринтов) лишено способности ориентироваться в пространстве при выключении зрения. Такое животное самостоятельно не пойдет в то место, куда его отводили за пищей не только однажды, а десятки раз. Этим самым доказывается, что в этом пищевом поведении кожно-мышечные и обонятельные ощущения заметной роли не играют.
Лабиринтные восприятия принимают самое активное участие в ориентации в пространстве даже при открытых глазах. Это видно из того, что безлабиринтные животные с открытыми глазами в вышеприведенных опытах поступают заметно иначе. Если безлабиринтное животное отнести с открытыми глазами в какое-либо место экспериментальной комнаты или в соседнюю комнату и там показать пищу, а затем отвести назад в исходное место, оно потом (при открытых глазах) не бежит туда кратчайшим путем, а перебегает от одного знакомого пищевого места к другому и так достигает показанного места пищи. Следовательно, прямолинейное ориентированное передвижение животного к пищевой цели по образу ее местонахождения существенно зависит от деятельности лабиринтных рецепторов. Именно когда нормальное животное с открытыми глазами передвигается по короткому пути к необычному месту за пищей, у него возникает образ этого пути, приобретенный на основании зрительной и лабиринтной рецепции, которым оно и руководствуется в дальнейшем при передвижениях к этому месту (Беритов, 1,5). Когда нормальное животное проходит многократно один и тот же путь к месту пищи, данное пищевое поведение автоматизируется. Оно начинает протекать по принципу цепного условного рефлекса, где каждый отрезок поведения вызывается через временные связи, образовавшиеся в ответ на те раздражения кожных, мышечных, слуховых и лабиринтных рецепторов,
которые происходили до этого отрезка поведения. В этом автоматизированном поведении как зрительные ощущения или зрительные образы, так и лабиринтные ощущения или пространственные образы играют существенную роль. Это видно из того, что если закрыть нормальному животному глаза и применить сигнал к данному пищевому поведению, то оно может побежать точно к месту пищи. Если же разрушить лабиринты и затем испытать условный сигнал (кожно-мышечное раздражение от сотрясения клетки) к автоматизированному пищевому поведению, собака с закрытыми глазами выйдет и& клетки, будет производить искательные движения по комнате, но прямо к месту пищи не пойдет. Если ей открыть глаза, она может побежать к месту пищи прямо без искательных движений, как при нормальных лабиринтах. Очевидно, зрительные восприятия от экспериментальной обстановки действуют как условные сигналы к данному пищевому поведению или к какомулибо отрезку его.
У безлабиринтного животного можно также образовать автоматизированное пищевое поведение по одному определенному пути даже при таких сложных условиях, как посещение трех кормушек в одном и том же порядке. Однако у безлабиринтной собаки при завязанных глазах для этого требуется не менее сотни подкармливают из данных кормушек, в то время как у нормальных животных достаточно было бы около десятка.
Из этих фактов следует, что автоматизированное пищевое поведение у нормальных животных осуществляется деятельностью временных связей, которые образуются главным образом на зрительные и лабиринтные раздражения. При разрушенных лабиринтах и закрытых глазах автоматизация поведения, очевидно, осуществляется главным образом деятельностью временных связей, образуемых на проприорецептивные раздражения.
Дата добавления: 2017-01-13; просмотров: 914;