Воспринимающая область обоняния.
Первичный нервный путь обонятельных рецепторов, расположенных в обонятельном эпителии, оканчивается в обонятельной луковице. Сюда притекают афферентные импульсы возбуждения по обонятельным нитям, которые фактически являются аксонами чувствительных обонятельных клеток. Эти импульсы активируют в обонятельной луковице митральные клетки, с которыми обонятельные нити связаны синаптически (рис. 56 — /).
Обонятельная луковица, являющаяся древним филогенетическим образованием, особенно развита у животных, в жизни которых обонятельная функция играет наиболее важную биологическую роль — в добывании пищи, в обнаружении жизненно опасных объектов, в сексуальных рефлексах.
Аксоны митральных клеток обонятельной луковицы входят в состав обонятельного тракта и направляются в разные отделы головного мозга. Морфо-
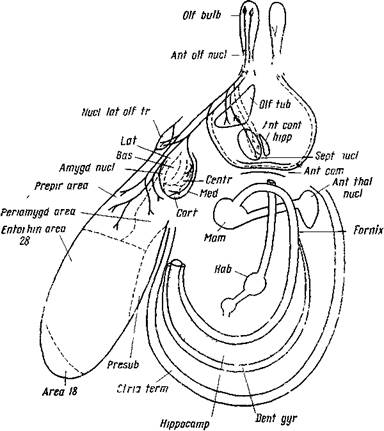
Рис. 56—Л. Схема связей обонятельной луковицы с разными отделами головного мозга
логическими работами, особенно Район Нахала (1911), было показано, что волокна обонятельного тракта заканчиваются главным образом в грушевидной извилине и миндалине. В настоящее время, особенно после работ Кларка и Мейера (124), известно, что эти волокна входят в обонятельный бугорок, в передний отрезок грушевидной извилины (препириформная область) и в комплекс миндалины (рис. 56—//).
Уайт, исследовавший корковую проекцию обонятельной луковицы у крыс, отмечает, что нейронная организация и система восходящих путей позволяют при минимальной активации обонятельной луковицы вызывать значительное возбуждение в обонятельной коре (Уайт, 125).
Здесь следует отметить, что обонятельный путь не проходит через таламус: обонятельная функция является единственной сенсорной функцией, которая не имеет представительства в таламусе.
Электрофизиологическими исследованиями было показано, что импульсы из митральных клеток обонятельной луковицы, кроме грушевидной извилины и амигдального комплекса, направляются и в другие отделы риненцефалона (Бродал, 126; Бун-Иши Хасама, 127; Аллен, 128; Эдриан, 129). Исследования в этом направлении, особенно за последнее десятилетие, показали, что имеется представительство обонятельной функции в старой коре головного мозга (Венцель, 130).
Типичные первичные ответы были зарегистрированы у собаки на различные запаховые раздражения (запах ксилола, гвоздики, асафетиды) в передней (ростральной) части грушевидной извилины (Аллен, 128). Эти
ответы возникали и при одиночных раздражениях обонятельной луковицы.
При раздражении той части грушевидной извилины, где возникали первичные ответы, в обонятельной луковице регистрировались ответные потенциалы. Этот факт, безусловно, указывает на наличие двусторонней нервной связи между обонятельной луковицей и определенными пунктами ростральной части грушевидной извилины.
При раздражении обонятельной луковицы у кошки наблюдались вызванные потенциалы в более обширных областях ростральной части грушевидной извилины, а также в периамигдаллярной области и с поверхности обонятельного тракта (Роуз и Булей, 131). Удалось зарегистрировать вызванные потенциалы еще из обонятельного бугорка, головки хвостатого ядра вентрально, из шелухи (путамен) бледного шара и энтопедункулярного ядра (Фоке, Мак Кинли и Мэгун, 132). Но не удалось получить при обонятельных раздражениях определенных ответов в гиппокампе и в миндалевидном комплексе. На отсутствие «обонятельных» вызванных потенциалов в гиппокампе указывают и другие авторы (133). На основании этих работ можно заключить, что, несмотря на анатомические данные о связи между обонятельной луковицей и миндалевидным комплексом, о чем говорилось выше, этот комплекс функционально не должен быть причислен к субстрату, первично воспринимающему обонятельную информацию.
Из грушевидной Р1звилины импульсы возбуждения обычно иррадиируют в различные области, в том числе и в гиппокамп. Может быть, факт наличия анатомической связи между грушевидной извилиной и гиппокампом был причиной тому, что до последнего времени гиппокамп часто рассматривался как образование, играющее важную роль в обонятельной функции. Однако наибольшее количество афферентных волокон идет в гиппокамп из каудальной, а не ростральной части грушевидной извилины. При раздражении обонятельной луковицы каудальная часть этой извилины (энторинальная область) или полностью молчит, или же дает низковольтные ответы с очень большой латентностью, что указывает на то, что эти потенциалы возникают в результате активации множества синаптических включений. На самом деле анатомические данные Кахала и Лоренте де Но говорят о том, что именно энторинальная область отдает волокна в гиппокапи. Эти данные в последнее время были подтверждены Алленом (128), который применил метод дегенерации волокон и показал, что после удаления энторинальной области наступает дегенерация волокон, идущих в гиппокамп, а при удалении ростральной части грушевидной извилины это не наблюдается.
Что гиппокамп нельзя считать первичной воспринимающей областью для обонятельных раздражений, видно, между прочим, из опытов Аллена (134), который показал, что после двустороннего удаления гиппокампа, так же как и затылочных, теменных и височных областей коры, у кошки не нарушаются условные рефлексы, выработанные на обонятельные раздражения. С другой стороны, хорошо известно, что в гиппокампе конвергируют афферентные импульсы от различных экстерорецепторов и интерорецепторов (Гриин и Ардуини, 135; Гриин и Махне, 136; Дзидзишвили, 137, 138; Унгиадзе, 139, и др.). Трудно себе представить, почему обонятельные раздражения должны составлять в этом отношении исключение? Известно, что обонятельная функция, которая филогенетически отмечена на самых ранних ступенях (Эдингер, 140), сохраняется в виде определенных реакций даже после удаления новой коры у высших позвоночных (Дюссер де Баренн, 141 и др.). Так, например, кошки с удаленным неокортексом находят пищу по запаху (Орджоникидзе и Нуцубидзе, 47).
По современным данным, гиппокамп играет важную роль в производстве общих эмоциональных реакций. Можно думать, что при обонятельных раздражениях гиппокамп играет также роль в производстве эмоциональных реакций при различных поведенческих актах. Эти реакции наступают в результате того, что при возбуждении обонятельных рецепторов информация
поступает вторично и в гиппокамп, наряду с другими структурами. В результате этого и наступают такие реакции, как сексуальное возбуждение на запахи, состояние тошноты, чувство отвращения или, наоборот, эмоция удовольствия от определенных запахов и т. п. Все эти реакции, наступающие вторично от активирования гиппокампа, и производят впечатление непосредственного участия гиппокампа в восприятии обонятельной функции.
Что касается эфферентных связей в обонятельном рецепторе, то имеются данные о наличии центрифугальных волокон вплоть до гранулярного слоя (Крэг, 142; Ломан и др., 143; Поуэлл, Коуен и Райзман, 144). Показано также влияние антидромных раздражений на обонятельные разряды. Кэрр и Хагбарт (145) нашли, что раздражение передней комиссуры тормозит эти разряды, тогда как раздражение латерального тракта не эффективно. Перерезка передней комиссуры снимает влияния, наблюдаемые при раздражении контралатерального обонятельного тракта (Манциа, Баумгартен и Гриин, 146). Эти авторы также показали, что раздражение ретикулярной формации приводит обычно к торможению одних нейронов и к активированию некоторых других нейронов обонятельной луковицы.
Дата добавления: 2017-01-13; просмотров: 794;