Воспринимающая область лабиринтных раздражений.
Волокна вестибулярного нерва оканчиваются на своей же стороне в вестибулярных ядрах продолговатого мозга. Отсюда вторичный путь частью поднимается в промежуточный мозг и далее в кору, а частью входит в сетевидное образование продолговатого и промежуточного мозга и через них также действует на кору (Бродал и др., 73).
Как указывалось выше, от раздражения рецепторов полукружных каналов и преддверия зависит, с одной стороны, восприятие изменений положения головы в пространстве, а с другой — восприятие пройденного пути (Беритов, 74). В коре большого мозга собаки и кролика восприятие лабиринтных раздражений осуществляется в основном передней частью эктосильвиевых
|
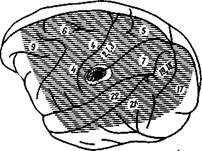
Рис. 53—I. Полушарие большого мозга
Масаса mulatto,
Горизонтальными линиями обозначается топографическое распространение вызванных потенциалов на раздражение вестибулярного нерва противоположной стороны. В центре рисунш первичная корковая проекция вести, булярного нерва в передней части по. стцентральной извилины в нижнеч конце интрапариетальной борозды. Цифры приблизительно обозначают поля по Бродману (по Фредриксон и др.)
и суправильвиевых извилин. У собак эта область перекрывает сзади слуховую область, а спереди — область кожной чувствительности. С помощью отведения вызванных потенциалов была определена локализация первичной области представительства вестибулярного нерва в коре. Так, у обезьян она локализована в задней части постцентральной извилины у основания интрапариетальной извилины между первым и вторым соматосенсорными полями, возможно, в поле 2, по Бродману. Это поле частично перекрывается первичной соматической областью, отвечающей на раздражение контралатерального срединного нерва, причем имеется взаимодействие между афферентами вестибулярного и срединного нервов в зоне перекрытия в виде конвергенции импульсов от этих нервов на некоторые нейроны вестибулярной области.
У обезьян вестибулярные ответы проявляются почти по всей поверхности новой коры. Первичная зона, возбуждаемая по специфической системе, занимает совсем небольшой участок (рис. 53—/). У кошек эта область также обширная, но заметна меньше (рис. 49—II). Эти ответы отличаются от наступающих в первичной зоне: они наступают через неспецифическую систему с большим скрытым периодом (30—400 мсек.), в то время как в первичной зоне скрытый период их 4—25 мсек. (Юнг и сотр., 36; Фредриксон и сотр., 75).
Топография распространения вестибулярных ответов устанавливалась путем осциллографической регистрации электрической активности коры (Кемпинский, 76; Вольц и Маунткэстл, 77; Майкл и Адес, 78). Однако первичные ответы при раздражениях лабиринтных рецепторов и вестибулярного нерва наблюдаются и по всей эктосильвиевой извилине (Андерсон и Гернанд, 79) и даже в зрительной коре (Юнг, 36; Горгиладзе, 80). Ответы на вестибулярные раздражения были найдены также при отведении активности нейронов в моторной коре (Корнхюбер и Ашофф, 81) (рис. 53—I).
Топография воспринимающей области лабиринтных раздражений устанавливалась и путем экстирпации коры больших полушарий. Было найдено, что наиболее устойчивые расстройства восприятий лабиринтной рецепции наблюдались при повреждении передней половины эктосильвиевых извилин, т. е.
если повреждается означенная первичная зона. Они выражались в том, что животные не воспринимали пройденного расстояния. Если нормальную собаку или кошку отвести с закрытыми глазами в необычное место на несколько метров и там дать поесть или понюхать пищу, потом она сама пойдет туда точно к месту пищи, и притом не только с открытыми глазами, но и с закрытыми. Так случится даже в том случае, если собаку с завязанными глазами не отводить к кормушке на ногах за ошейник, а перенести в клетке в лежачем положении и после еды возвратить обратно в клетке тем же или другим путем. В этом случае собака также может пойти коротким путем к месту пищи. И вот, после экстирпации передней половины эктосильвиевых извилин, когда повреждается центральная первичная зона вестибулярной области, животное перестает воспринимать пройденное расстояние и теряет способность повторять пройденный путь (Беритов, 62).
Афферентный путь от лабиринтных рецепторов, как и от слуховых рецепторов, проходит не только прямо по стволу головного мозга, но и сложным путем, через мозжечок. Здесь лабиринтные афферентные волокна заканчиваются в задних долях (nodulus, flocculi, uvula) и в передней доле (lingula). Отсюда начинается новый афферентный путь, который кончается в коре мозга там же, где кончается прямой путь — в основном в передней части сильвиевых извилин. Это видно из того, что раздражение этих участков мозжечка вызывает электрические потенциалы главным образом в передней половине эктосильвиевых извилин (Руолдт и Снайдер, 82).
Разрушение означенных долей мозжечка, как и удаление всего мозжечка, влечет за собой долговременное нарушение восприятия пройденного пути на основе лабиринтной рецепции. Лишь спустя много месяцев после операции, наблюдается более или менее значительное восстановление способности к восприятию и проецированию пройденного пути.
У безмозжечковых животных не исчезает возможность сохранения равновесия при изменениях положения головы. Такие животные свободно передвигаются и на всякие изменения положения головы реагируют соответствующим тоническим рефлексом, тем самым сохраняя тело в равновесии. Следовательно, мозжечок играет важную роль в деятельности вестибулярной воспринимающей области, главным образом в отношении восприятия и проецирования пройденного пути.
Вопрос о центробежной эфферентной регуляции вестибулярного рецептора предстоит еще экспериментально обосновать. Он доказан косвенно, гистохимическими методами (Долман с сотр., 83) по локализации холинэстеразы в вестибулярном эпителии. Отчетливые результаты были получены для вестибулярной функции у лягушки (Глейснер и Хенриксон, 84). Электрофизиологическая регистрация эфферентной импульсации проведена на кроликах (Бертранд и Веенгоф, 85). Новое подтверждение этого положения получено в недавней работе Прехта с сотр. (Маркхам, Прехт, Шимацу, 86).
Дата добавления: 2017-01-13; просмотров: 684;