А. Поверхневі м’язи
1. зовнішній м’яз – замикач відхідника (m. sphincter ani externus). Цей м’яз у вигляді кільця оточує відхідник і прилеглу частину прямої кишки. Biн має три частини: підшкірну (pars subcutanea) (волокна якої оточують пряму кишку з боків, вплітаються в шкіру спереду i ззаду від відхідника); поверхневу (pars superfidalis), що починається від відхідниково-куприкової зв’язки (lig. anococcygeum) і верхівки куприка, обходить пряму кишку з боків i закінчується в промежинному тілі; глибоку (pars profunda) найпотужнішу – що у вигляді кільця оточує пряму кишку. М’яз переважно перебуває в скороченому стані, має посмуговані, м’язові волокна, що розслаблюються під час акту дефекації.
Б. Глибокі м’язи
1. м’яз-підіймач відхідника (m. levator ani) парний (правий та лівий), закриває вихід із малого таза. М’яз починається від нижньої гілки лобкової кістки, затульної фасції і тазової поверхні сідничної кістки. Пучки волокон м’яза проходять збоку від прямої кишки, обходять її ззаду, утворюючи з протилежним м’язом петлю. У чоловіків м’яз проходить повз передміхурову залозу, з’єднуючись з нею м’язовими волокнами. М’яз прикріплюється до верхівки та бічних країв куприка, до відхідниково-куприкової зв’язки. Функція: піднімає дно таза і відхідникову частину прямої кишки, притискає задню стінку прямої кишки до її передньої стінки.
2. лобково-куприковий м’яз (m. coccygeus) рудиментарний. Починається від сідничої ості та крижово-остьової зв’язки, прикріплюється до краю куприка i верхівки крижової кістки. Функція: зміцнює тазову діафрагму.
Фасції промежини.
До них належать: фасція промежини, нижня і верхня фасція тазової діафрагми.
Фасція промежини (fascia perinei) розташовується глибше від підшкірної жирової клітковини промежини, має вигляд тонкого прошарку. У чоловіків фасція продовжується в м’ясисту оболонку калитки та у фасцію статевого члена. Фасція промежини покриває поверхневий поперечний м’яз промежини. У відхідниковій ділянці фасція виділяє підшкірну жирову клітковину від жирового тіла сіднично-відхідникової ямки.
Нижня фасція тазової діафрагми (fascia inferior diaphragmatis pelvis). Розгляд її почніть з вихідникової ділянки, де вона є продовженням фасції, що покриває великий сідничний м’яз. Продовжуючись вперед від нижнього краю великого сідничного м’яза, нижня фасція тазової діафрагми переходить на нижню поверхню м’яза-підіймача ви хідника. Дійшовши до поверхневого поперечного м’яза промежини, фасція поділяється на три шари - поверхневий, середній та глибокий. Поверхневий шар покриває поверхневий поперечний м’яз промежини, сідничо-печеристий та цибулинно-печеристий м’язи. Середній шар покриває нижню поверхню глибокого поперечного м’яза промежини і зовнішнього м’яза-замикача сечівника. Глибокий шар покриває верхню поверхню глибокого поперечного м’яза промежини і зовнішнього м’яза-замикача сечівника.
Поверхневий та середній шар нижньої фасції тазової діафрагми зростаються з нижніми гілками лобкових кісток та з гілками сідничних кісток, між ними розташовані печеристі тіла статевого члена (ніжки клітора).
Окрім того середній та глибокий шари у місці з’єднання з нижніми гілками лобкових кісток з’єднуються ще й між собою, утворюючи перетинку промежини. У щілині між нижньою лобковою зв’язкою i перетинкою промежини проходять спинкові артерія і вена статевого члена (клітора).
Верхня фасція тазової діафрагми (fascia superior diaphragmatis pelvis) у ділянці тазової діафрагми покриває верхню поверхню м’яза-підіймача відхідника. Дійшовши до рівня заднього краю глибокого поперечного м’яза промежини, фасція з’єднується з глибоким шаром нижньої фасції діафрагми таза.
Сідничо-відхідникова ямка (fossa ischioanalis) обмежована:
з боків - внутрішній затульний м’яз, покритий затульною фасцією, i присередня поверхня сідничого горба;
медіально - нижньою поверхнею м’яза-підіймача відхідника і зовнішнім м’язом-замикачем відхідника;
ззаду - задніми пучками м’яза-підіймача відхідника i сідничо-куприковим м’язом;
спереду – поверхневим та глибоким поперечним м’язами промежини.
З обох боків від прямої кишки в сідничо-відхідниковій ямці розміщується жирове тіло сідничо-відхідникової ямки (corpus adiposum fossae ischiorectalis). Тут можуть розвиватися запальні процеси та нагноєння жирової клітковини.
Особливості м’язів зовнішніх статевих органів жіночої промежини.
Цибулинно-губчастий м’яз проходить справа та зліва від піхви. Він починається від промежинного тіла, вкриває з боку велику присінкову залозу, цибулину присінка і вплітається в печеристе тіло клітора. Функція: стискує вени, що відводять кров з цибулини присінка, сприяючи наповненню її кров’ю; звужує отвір піхви; стискує великі залози присінка, сприяючи виділенню з них секрету.
Сідничо-печеристий м’яз прикріплюється до білкової оболонки печеристих тіл клітора на його спинці. Функція: стискує вени і печеристі тіла клітора, сприяючи просуванню крові до головки клітора i перешкоджає відтіканню її у вени.
Зовнішній м’яз - замикач сечівника у жінок охоплює не тільки сечівник, а й піхву. Пучки м’яза обходять з боків сечівник та піхву i спереду від сечівника перехрещуються з волокнами протилежного м’яза, закінчуючись у сполучній тканині між сечівником і перетинкою промежини. Позаду від піхви волокна м’яза закінчуються в промежинному тілі. Функція: стискає сечівник i отвір піхви.
Особливістю м’яза - підіймача відхідника у жінок є те, що м’яз проходить повз піхву та сечовий мixyp, вплітається м’язовими й еластичними волокнами в їx стінки. Завдяки скороченню притягає задню стінку піхви до передньої, звужуючи отвір піхви, піднімає дно таза та відхідникову частину прямої кишки.
Волокна глибокого поперечного м’яза промежини у жінок з’єднуються зі стінкою піхви, в його товщі розташовані великі залози присінка. Функція: скорочуючись, він стискає великі залози присінка, сприяючи виділенню з них секрету.
Залози внутрішньої секреції
(glandulae endocrinae)
Регуляція функцій організму людини здійснюється крім нервової системи гормонами ендокринних залоз. В той же час ендокринна, або гуморальна (хімічна) регуляція підпорядкована нервовій системі. Обидві системи функціонально найтіснішим чином між собою взаємодіють і становлять єдину нейрогуморальну регуляцію організму. Крім того, залози внутрішньої секреції знаходяться в дуже складних взаємовідносинах між собою, так що порушення роботи однієї з них відображається на діяльності інших. Хімічна регуляція діє протягом тривалого часу, стимулюючи або пригнічуючи процеси обміну речовин і в самій нервовій системі.
Залози внутрішньої секреції за їх ембріональним розвитком поділяються на такі групи:
1. ектодермальні залози:
а) гіпофіз і шишкоподібна залоза, що розвиваються з епітелію ротової бухти та проміжного мозку;
б) мозкова речовина надниркових залоз, що розвиваються з нервової трубки;
2. мезодермальні залози:
а) кіркова речовина надниркових залоз;
б) ендокринна частина яєчка або яєчника;
в) ендокринна частина передміхурової залози, що розвиваються з епітелію вторинної порожнини тіла;
3. ентодермальні залози:
а) бранхіогенна група, що розвивається з епітелію ротової бухти, зябрових кишень (щитоподібна залоза; прищитоподібні залози, загруднинна залоза);
б) ендокринна частина підшлункової залози (острівці підшлункової залози, що розвиваються з епітелію кишкової трубки).
За функціональною взаємозалежністю залози внутрішньої секреції поділяються на такі групи:
1. Група аденогіпофіза:
а) щитоподібна залоза;
б) кора надниркових залоз (пучкова та сітчаста зони);
в) яєчко чи яєчник.
Основна роль у цій групі залоз належить аденогіпофізу, його гормонам (аденокортикотропному, соматотропному, тіреотропному і гонадотропному), які регулюють діяльність названих залоз.
2. Група периферійних ендокринних залоз, функція яких не залежить від гормонів аденогіпофіза:
а) прищитоподібні залози;
б) кора надниркових залоз (клубочкова зона);
в) острівці підшлункових залоз.
Ці залози умовно називають саморегулівними, наприклад, гормон острівців підшлункової залози інсулін знижує рівень глюкози в крові, а підвищений рівень останньої стимулює секрецію інсуліну.
3. Група ендокринних залоз нервового походження:
а) великі та малі нейросекреторні клітини з відростками, що утворені ядрами гіпоталамуса;
б) нейроендокринні клітини, які не мають відростків (хроматофінні клітини мозкової частини надниркових залоз та парагангліїв);
в) парафолікулярні клітини, або клітини щитоподібної залози;
г) аргірофільні та ентерохромафінні клітини в стінках шлунка, тонкої та товстої кишок.
Нейросекреторні клітини поєднують нервову та ендокринну функцію (характеристику нейросектеторної діяльності клітин гіпоталамуса викладено в темі “Проміжний мозок”). Вони сприймають нервові імпульси і виробляють у відповідь нейросекрет, який потрапляє в кров або по відросткам нервових клітин транспортується до інших клітин. Так, клітини гіпоталамуса продукують нейросекрет, який по відростках нервових клітин надходить у гіпофіз і підсилює або гальмує його функцію.
4. Група ендокринних залоз нейрогліального походження:
а) шишкоподібна залоза;
б) нейрогемальні залози (нейрогіпофіз та серединне підвищення гіпоталамуса).
Секрет, що виробляється клітинами шишкоподібного тіла, гальмує виділення гонадотропних гормонів аденогіпофіза і, таким чином, пригнічує діяльність статевих залоз. Клітини задньої частки гіпофіза не продукують гормонів, а забезпечують накопичення і виділення в кров вазопресину та окситоцину, що продукуються клітинами гіпоталамуса.
Щитоподібна залоза (glandula thyroidea) - непарний орган масою 20-50 г; у віці після 55-60 років об’єм та маса залози зменшується.
Залоза розташована в передній ділянці шиї на рівні гортані та верхнього відділу трахеї; складається з двох часток - правої частки (lobus dexter)та лівої частки (lobus sinister), з’єднаних вузьким перешийком щитоподібної залози (istmus glandulae thyroidei), який розташований на рівні II-III хрящів трахеї (мал. 233). Якщо перешийок відсутній, частки залози роз’єднані. Верхні краї правої та лівої часток щитоподібної залози розташовані нижче від верхнього краю відповідної пластинки щитоподібного хряща гортані. Нижні краї обох часток досягають рівня V-VI хрящів трахеї. Задня поверхня кожної частки залози прилягає до гортанного відділу глотки, початкового відділу стравоходу, а також межує з передньою поверхнею загальної сонної артерії. У 30% випадків зустрічається пірамідна частка (lobus pyramidalis), яка верхівкою іноді досягає тіла під’язикової кістки.
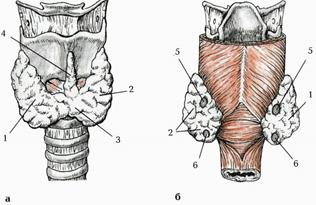
Мал. 222. Щитоподібна залоза. А - вигляд спереду, Б - вигляд ззаду. 1. Lobus dexter. 2. Lobus sinister. 3. Isthmus glandulae thyroideae. 4. Lobus pyramidalis. 5. Glandula parathyroidea superior. 6. Glandula parathyroidea inferior.
Щитоподібна залоза покрита сполучнотканинною оболонкою - волокнистою капсулою (capsula fibrosa), яка зрощена з гортанню та трахеєю. У зв’язку з цим під час акту ковтання відбувається зміщення щитоподібної залози. У товщу залози від капсули відходять сполучнотканинні перетинки, які поділяють її тканину на часточки (lobuli). Ці часточки вистелені зсередини епітеліальними фолікулярними клітинами кубічної форми.
Структурно-функціональною одиницею щитоподібної залози є фолікул - утворення кубічної форми, виповнене густою речовиною (колоїдом). Остання містить гормон щитоподібної залози тироксин, до складу якого входять білки й амінокислоти, що адсорбують йод. Активна форма гормону - тироксин, або Т4 - тетрайодтиронін. Зменшення його кількості в крові збільшує вихід тиреотропного гормону гіпофіза (ТТГ), який в свою чергу стимулює функцію щитоподібної залози, тобто підвищує синтез тироксину за механізмом від’ємного зворотного зв’язку. Головний ефект дії тироксину полягає в підвищенні споживання кисню більшістю органів і тканин людини. У разі збільшення споживання організмом кисню і виділення тепла в ньому відбувається посилений розпад білків і жирів. Якщо рівень гормонів щитоподібної залози підвищується (гіпертиреоз), розвивається дифузний токсичний зоб (базедова хвороба). Гіпертиреоз супроводжується підвищенням секреції гормонів залози, симптомами якого є нервозність, тремтіння рук, тахікардія, пітливість, втома, втрата маси тіла, екзофтальм, збільшення розмірів щитоподібної залози. Синтез гормонів при цьому збільшується в 3-10 разів.
У разі зниження функції щитоподібної залози зменшується споживання кисню та знижується швидкість обмінних процесів, розвивається гіпотиреоз. Якщо гіпофункція щитоподібної залози набуває стійкого характеру, у дітей розвивається кретинізм; у дорослих мікседема (набряки шкіри), знижується основний обмін.
Прищитоподібні залози парні (дві верхні та дві нижні). Верхня прищитоподібна залоза (glandula parathyroidea superior) і нижня прищитоподібна залоза (glandula parathyroidea inferior)належать до числа життєво необхідних органів (див. мал. 233 б). Розміри кожної залози такі: довжина 4-8 мм, ширина 3-4 мм, товщина 2-3 мм. Загальна маса залоз становить у середньому близько 1 г. У дітей залоза блідо-рожевого кольору, у дорослих – жовто-коричневого. Залози мають власні волокнисті капсули, добре кровопостачаються. Прищитоподібні залози виділяють паратгормон (ПтГ), що регулює рівень кальцію в крові та м’яких тканинах. Низький вміст кальцію в крові посилює викид ПтГ в кровоносне русло. ПтГ активує остеокласти, які руйнують кісткову речовину, сприяючи її резорбції, при цьому у кровоносне русло надходить вільний кальцій. Органічна речовина кісток розпадається і разом з кальцієм виводиться через нирки. Підвищена кількість кальцію в крові може спричинятися до утворення каменів у сечових шляхах.
Надниркова залоза (glandula suprarenalis) - парний ендокринний орган, розміщений над верхнім кінцем нирки в заочеревинному просторі (мал. 234). Розрізняють три поверхні залози: передню (facies anterior), задню (facies posterior) і ниркову (facies renalis). Маса залози 12-13 г, довжина 5 см, ширина 3 см, висота 1 см. На передній поверхні розміщені ворота (hіlum), через які із залози виходить центральна вена (v.centralis). Зовні надниркові залози покриті сполучнотканинною оболонкою, до якої прилягає кіркова речовина (cortex). Вона складається з трьох зон: клубочкової (zona glomerulosa), за нею розміщена найширша пучкова зона (zona fasciculusa), а на межі з мозковою речовиною надниркової залози (medulla)- сітчаста зона (zona reticularis). Ширина цих зон у товщі кіркової речовини співвідноситься, відповідно, 1:9:3.
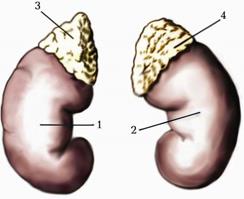
Мал. 223. Права і ліва нирки з наднирковими залозами. 1. Ren dexter. 2. Ren sinister. 3. Glandula suprarenalis dextra. 4. Glandula suprarenalis sinistra.
Великі клітини ендокриноцити пучкової зони розміщаються рядами (“пучками”). Ендокриноцити синтезують глюкокортикостероїдні гормони (кортизон, гідрокортизон, кортикостерон), які регулюють обмін вуглеводів, білків, ліпідів, стимулюють енергетичний обмін, а також пригнічують запальні процеси в організмі.
Ендокриноцити сітчастої зони синтезують статеві гормони, більшою мірою чоловічі стероїди, подібні до тестостерону яєчок, меншою мірою - жіночі статеві гормони естрогени і прогестерон.
Кора надниркових залоз виробляє кортикостероїди, які поділяються на три групи: а) мінералокортикостероїди (альдостерон) – їх продукує клубочкова зона; б) глюкокортикостероїди, до яких крім кортизону, гідрокортизону, кортикостерону, належать ще два гормони – II-дегідрокортикостерон і II-дезоксикортикостерон, які виробляються в пучковій зоні; в) статеві гормони (андроген, естроген, прогестерон), які виробляються в сітчастій зоні.
Мозкова речовина надниркової залози, що розташована в її центрі, побудована з великих клітин округлої або полігональної форми, які виробляють адреналін і норадреналін. Ці гормони називають катехоламінами, вони мобілізують захисні сили організму. Завдяки підвищенню рівня гормонів у крові розщеплюється глікоген, зменшується його запаси в м’язах та печінці.
Кіркова і мозкова речовини надниркової залози мають різне походження. Кіркова речовина розвивається з мезодерми (целомічного епітелію) між коренем дорсальної брижі первинної кишки і сечово-статевою складкою. Мезодермальні клітини розташовуються між двома первинними утвореннями нирок і називаються інтерреналовою тканиною. Вони дають початок кірковій речовині, іноді з неї утворюються додаткові надниркові залози (glandulae suprarenalis accessoriae).
Мозкова речовина надниркових залоз має спільне з нервовою системою походження. Вона розвивається з ембріональних нервових клітин симпатобластів, які формуються із закладок вузлів симпатичного стовбура, і відокремлюються від нього, перетворюючись на хромафінобласти, а останні - на хромафінні клітини мозкової речовини.
Хромафінобласти мігрують з вузлів симпатичного стовбура й утворюють поза хромафінні тільця - розташовані вздовж черевної частини аорти - парні аортальні параганглії (paragangla aortica), у товщі вузлів симпатичного стовбура - симпатичні параганглії (paragangliа sympathicа).
Вивчить функцію надниркових залоз, яка тісно пов’язана з ендокринною функцією нирок. Відомі дві основні гормональні системи нирок - ренінангіотензивна система, що тісно пов’язана з функцією надниркових залоз, печінки, легень, та система простагландинового комплексу.
У нирці виробляється і виділяється в кров гормон ренін. Він синтезується в спеціальній групі епітеліальних клітин, названих юкстагломерулярними. Ці клітини вистеляють приносну артеріолу і розташовані між нею та частиною дистального канальця (мал. 235). Різні чинники (зокрема, зниження ниркового перфузійного тиску, посилення активності симпатичного відділу автономної нервової системи, зменшення загального об’єму позаклітинної рідини в організмі, зміна електролітного складу рідини в дистальному канальці, до яких чутливі юкстагломерулярні клітини, стимулюють вихід реніну.
Ренін діє на глобулін плазми крові, що синтезується в печінці та утворює ангіотензин-І. Під впливом особливого ферменту, який знайдено в легенях та нирці, ангіотензин-I розщеплюється, утворюючи ангіотензин-II. Біологічно активний ангіотензин-II є судинозвужувальним агентом. Таким чином, ренінангіотензивна система бере участь у виникненні та розвитку гіпертонічної хвороби.
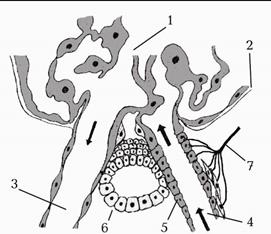
Мал. 224. Схема юкстагломерулярного апарату нирки.
1. Клубочок.
2. Інтерстицій.
3. Еферентна артеріола.
4. Аферентна артеріола.
5. Юкстагломерулярні клітини.
6. Macula densa.
7. Нерви.
Другим основним ендокринним комплексом у нирці є простагландинова гормоноподібна система, яка крім нирок впливає на серцево-судинну систему і нормалізує її діяльність.
Наднирникові залози продукують дві групи гормонів: гормони кори надниркових залоз і гормони мозкової речовини.
Ендокриноцити клубочкової зони кори надниркових залоз виділяють мінералокортикостероїдний гормон альдостерон, який регулює обмін натрію, беручи участь у підтриманні загального гомеостазу й водно-сольового обміну. Разом з реніном, альдостерон бере участь у розвитку тяжкої хвороби артеріальної гіпертонії. Підвищений вміст у крові альдостерону і реніну призводить до виникнення артеріальної гіпертензії. Крім того, альдостерон має здатність посилювати перебіг запальних процесів.
У разі порушення функції кори надниркових залоз (атрофія, ушкодження) розвивається аддісонова хвороба - первинна недостатність надниркових залоз, для якої характерно зниження синтезу альдостерону і кортизону. Дефіцит цих гормонів і, передусім (альдостерону) призводить до зниження іонів натрію в крові і тканинах (гіпонатріємія) та підвищення рівня іонів калію (гіперкаліємія).
Зменшення кількості натрію супроводжується втратою тканинної рідини і зневодненням. Падіння рівня кортизону призводить до слабкості, сонливості. На шкірі у відкритих місцях з’являється відкладення пігменту меланіну, шкіра набуває бронзового кольору.
Острівці підшлункової залози (insulaе pancreaticі) відокремлені від екзокринної частини підшлункової залози тонкими сполучнотканинними прошарками (мал. 236). Острівців більше у хвості і менше в головці підшлункової залози. Величина острівців коливається від 0,1 до 0,3 мм і становить 1-3,6% маси всієї залози. Загальна кількість острівців у залозі від 200 тис. до 2 млн. Вони складаються з ендокринних клітин інсулоцитів, які розвиваються з ентодерми середнього відділу первинної кишки. Між клітинами локалізовані гемокапіляри фенестрованого типу, оточені перикапілярними просторами. Гормони потрапляють в пери капілярний простір, а з них через стінку капіляра у кров.
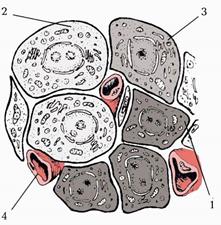
Мал. 225. Схема гістологічної будови острівця підшлункової залози.
1. — клітини.
2. — клітини.
3. — клітини.
4. Кровоносний капіляр.
Ендокринна частина статевих залоз -це у чоловіків і яєчник у жінок, які крім статевих клітин, виробляють і виділяють у кров статеві гормони, що впливають на формування вторинних статевих ознак.
Ендокринна функція яєчок здійснюється інтерстиціальними ендокриноцитами, розташованими в пухкій сполучній тканині між звивистими сім’яними трубочками, поряд з кровоносними та лімфатичними капілярами. Інтерстиціальні ендокриноцити яєчка виділяють чоловічій статевий гормон тестостерон.
Дія тестостерону зумовлює розвиток вторинних статевих ознак: огрубіння голосу, оволосіння в пахвових та пахвинній ділянках, ріст волосся на обличчі, збільшення розмірів зовнішніх статевих органів. Під його впливом змінюється соматична статура: плечі стають широкими, розвивається м’язова система.
Якщо яєчка з певних причин не опустилися в калитку, тобто залишились у черевній порожнині, де температура вища, ніж у калитці, у чоловіка може бути безпліддя.
Стан, зумовлений порушенням функції інтерстиціальних ендокриноцитів, або відсутність яєчок унаслідок природжених вад чи кастрації, називається євнухоїдизмом. Для осіб, які страждають на євнухоїдизм, характерні такі особливості будови тіла: вузькі плечі, погано розвинені м’язи, жіночі риси зовнішності, невеликі розміри статевих органів, високий голос, відсутність оволосіння на лобку, жіночий тип розташування підшкірної жирової клітковини.
У яєчнику синтезуються статеві гормони естроген і прогестерон. Естроген, або фолікулін, продукується зернистим шаром дозріваючих фолікулів, а також клітинами інтерстицію яєчника. Ріст фолікулів і активізація інтерстиціальних клітин відбувається під впливом фолікулостимувального гормону гіпофіза. На овуляцію й утворення жовтого тіла впливає лютеїнізуючий гормон, якому властива ендокринна функція - клітини його виробляють гормон прогестерон, останній підготовлює слизову оболонку матки до сприйняття заплідненої яйцеклітини, а також затримує розвиток фолікулів.
Менструація та овуляція належать до циклічних процесів жіночого організму. Під час постменструальної фази фолікули ростуть в яєчнику, дозрівають і виробляють естрогени, а після овуляції з фолікула, який лопнув, утворюється жовте тіло, що секретує прогестерон - це гормон, який сприяє збереженню цілісності слизової оболонки матки і збільшенню його товщини. З цього моменту починається передменструальна фаза, або фаза жовтого тіла. Вона триває до кінця 28-го дня. Потім цикл повторюється.
Слід зауважити, що естрогени виробляються яйцеклітиною і відіграють важливу роль у менструальному циклі, сприяючи розвитку матки та грудних залоз. У наслідок дії естрогенів у пубертатний період розвиваються вторинні статеві ознаки.
Гіпофіз (hypophysis) розташований в гіпофізній ямці турецького сідла клиноподібної кістки і відокремлений від загальної порожнини черепа твердою оболоною головного мозку у вигляді діафрагми сідла. Через отвір цієї діафрагми гіпофіз з’єднаний лійкою з сірим горбом гіпоталамуса проміжного мозку. Розміри гіпофіза у дорослої людини такі: вертикальний - 0,5-1 см; поперечний - 1-1,7 см; передньозадній - 0,5-1,6 см. Гіпофіз має форму квасолі, його маса у чоловіків 0,5 г, у жінок 0,6 г. Зовні гіпофіз покритий капсулою. Отвір діафрагми сідла над гіпофізом менший, ніж розміри порожнини гіпофізної ямки, у зв’язку з чим під час видалення із порожнини черепа півкуль головного мозку ніжка лійки гіпофіза відривається і він залишається в гіпофізній ямці турецького сідла.
Гіпофіз поділяється на дві частки - передню (lobus anterior) і задню (lobus posterior).
Передня частка називається аденогіпофізом (на його долю припадає 70-80% від загальної маси залози). Клітини аденогіпофіза синтезують та виділяють у кров тропні гормони, функція яких полягає у регуляції діяльності ряду периферійних ланок ендокринної системи, тобто гіпофізозалежних органів; а також впливає на лактоцити грудної залози, меланоцити, сперматогонії яєчка тощо.
Задня частка має назву нейрогіпофіза, у ній накопичуються та виділяються в кров синтезовані нейросекреторними клітинами переднього гіпоталамуса гормони окситоцин та вазопресин.
Зверніть увагу на механізм дії гормонів аденогіпофіза:
i тиреотропний гормон (ТТГ) стимулює ріст і дозрівання клітин щитоподібної залози, сприяє синтезу і вивільненню гормонів;
i адренокортикотропний гормон (АКТГ) стимулює ріст і дозрівання клітин кіркового шару надниркових залоз, синтез і вивільнення кортикостероїдів;
i фолікулостимулювальний гормон (ФСГ) стимулює ріст і розвиток фолікулів яєчника з наступним виходом з них естрогенів, а також ріст яєчок і сперматогенез;
i лютеїнізувальний гормон (ЛГ) спонукає до періодичного виходу яйцеклітини з яєчника (овуляції), а також наступного розвитку жовтого тіла, що секретує прогестерон; крім того ЛГ сприяє росту і розвитку інтерстиціальних клітин яєчка, які виробляють андрогени;
i соматотропний гормон (СТГ), сприяє росту і розвитку кісток та інших тканин у ранньому віці;
i меланостимулювальний гормон (МСГ) стимулює накопичення меланіну в шкірі;
i пролактин- головна його функція полягає в ініціації та підтриманні лактації.
Для фізичного розвитку людини важливе значення має соматотропний гормон (СТГ). У разі його недостатності в дитячому віці виникає карликовість (нанізм). У таких людей спостерігається зниження рівня й інших гормонів, а часом - затримка статевого розвитку. Якщо ж секреція СТГ у дітей підвищена виникає гігантизм. Зріст таких людей досягає 2,5 м за рахунок збільшення довгих трубчастих кісток кінцівок.
Гіперсекреція СТГ в похилому віці призводить до акромегалії, тобто до збільшення: носа, вушних раковин, губ, підборіддя, м’яких тканин обличчя та пальців кінцівок.
Задня частка гіпофіза, або нейрогіпофіз утворена розширеннями аксонів нейросекреторних клітин надзорового та пришлуночкового ядер гіпоталамуса, які продукують гормони вазопресин та окситоцин.
Гормон вазопресинвиявляє судинозвужувальну та антидіуретичну дію, за що й отримав назву антидіуретичного гормону (АДГ). Ще раз зверніть увагу на функцію задньої частки гіпофіза, тобто нейрогіпофіза, який виробляє антидіуретичний гормон (АДГ), або вазопресин та окситоцин. АДГ впливає на дистальну частину канальця нефрона та збиральні канальці нирок, посилюючи реабсорбцію води і знижуючи діурез. Це важливо для організму тварин, які живуть у пустелях. На секрецію АДГ впливає кількість води у крові і тканинах.
У разі патологічного зниження секреції вазопресину виникає нецукровий діабет, для якого характерно збільшення діурезу і вживання великої кількості води. Зверніть увагу на те, які патологічні стани виникають, якщо порушується функція аденогіпофіза.
Порушення виділення ФСГ та ЛТГ є причиною затримки статевого розвитку, а також порушень жирового обміну і передчасного статевого дозрівання.
Гіперплазія базофільних клітин передньої частки гіпофіза призводить до вироблення надлишку АКТГ, що в свою чергу спричинює гіперплазію кори надниркових залоз і гіперпродукцію альдостерону. Це призводить до непропорційного відкладення жиру у верхній частині тіла: обличчя стає круглим, місяцевоподібним, ціанотичним, у ділянці спини горб, шкіра тіла має розтяги у вигляді білих смуг; хворі слабкі, страждають на цукровий діабет, шкіра суха, статеві функції знижені, психіка порушена. Таку патологію називають первинним альдостеронізмом, або хворобою Іценка-Кушинга.
Окситоцин стимулює скоротливу функцію м’язової оболонки матки, утворення молока у грудній залозі під час лактації, а також впливає на підвищення тонусу непосмугованих м’язів травного тракту.
Шишкоподібна залоза (glandula pinealis) належить до проміжного мозку і розташована в поздовжній борозні між верхніми горбками пластинки покрівлі середнього мозку. Тіло шишкоподібної залози має овоїдну, або конічну форму. Маса шишкоподібної залози у дорослої людини становить близько 0,2 г, довжина, в середньому, 1,2 см, ширина 0,8 см, товщина 0,5 см. Зовнішня поверхня залози покрита волокнистою капсулою (capsula fibrosa), від якої в її товщу відокремлюються перетинки, що ділять залозу на часточки (lobuli). Кожна часточка шишкоподібної залози складається з двох видів клітин - нейросекреторних пінеалоцитів і гліоцитів. Пінеалоцити синтезують близько 40 регулівних пептидів, а також біологічноактивні аміни - серотонін і мелатонін. Останній має здатність пригнічувати секрецію гонадоліберину гіпоталамуса, сповільнюючи статеве дозрівання. Максимального розвитку шишкоподібна залоза досягає на 7-ому році життя, після чого починається її вікова інволюція.
Центральним нейросекреторним органом, який поєднує нервову і гуморальну регуляцію, є гіпоталамус (hypothalamus). Гіпоталамус займає близько від 5% маси мозку. Через гіпоталамус центральна нервова система здійснює контроль над регуляцією функцій внутрішніх органів. Прийнято вважати, що гіпоталамус - це сукупність вищих центрів функцій внутрішніх органів. Гіпоталамус складається із скупчень нервових клітин, які утворюють близько 30 пар ядер, що розміщені в основі головного мозку, навколо III шлуночка. Розрізняють дзьобове, дорсальне проміжне, бічне та заднє гіпоталамічне поля.
У дзьобовому гіпоталамічному полі (аreа hypothalamica rostralis) є такі основні пари ядер: над зорове (nucleus supraopticus) та пришлуночкове (nucleus paraventricularis). Клітини цих ядер виробляють гормони вазопресин та окситоцин, які по аксонах нейросекреторних клітин опускаються в задню частку гіпофіза, звідки виводяться у кровообіг аксовазальнами синапсами (мал. 237).
До середнього або проміжного гіпоталамуса (regio hypothalamica intermedia) відносять передньомедіальне та дорсоприсереднє підталамічні ядра (nuclei hypothalamici ventromedialis et dorsomedialis); дорзальне підталамічне ядро (nucleus hypothalamicus dorsalis); лійкове ядро (nucleus infundibularis); сірогорбове ядро (nucleus tuberalis).
Нейрогормони, які виробляються клітинами ядер гіпоталамуса носять загальну назву гормон-регулівних факторів. Вони становлять систему переключення впливу нервової системи на ендокринну. Серед цих факторів виділяють ліберини і статини. Перші стимулюють фізіологічні процеси, другі – пригнічують їх. Досліджено сім ліберинів і три статини.
Ядра гіпоталамуса досить тісно пов’язані аферентними й еферентними шляхами з гіпофізом, вони здійснюють регулівний вплив на нервові автономні та ендокринні функції організму.
Нейросекрет ядер гіпоталамуса здатний впливати на функцію залозистих клітин гіпофіза, посилюючи або пригнічуючи секрецію деяких його гормонів, які регулюють діяльність окремих залоз внутрішньої секреції. Відбувається подвійна або сходинкова регуляція ендокринних залоз.
Підсумовуючи дію гормонів, необхідно засвоїти, що виділення кожного тропного гормону гіпофіза регулюється одним із гормон-регулівних факторів гіпоталамуса. Винятком є ФСГ та ЛГ, виділення яких не регулюються факторами гіпоталамуса.
Крім того, вироблення гіпофізом пролактину, а також сомато- і меланотропного гормонів відбувається під дією двох пар гормон-регулівних факторів: пролактоліберіну та пролактостатину; соматоліберину та соматостатину; меланоліберину та меланостатину.
Регуляція всіх інших гормонів та нейросекреції гіпоталамуса відбувається за механізмом зворотного зв’язку.
Гормони, що циркулюють у крові, можуть діяти на гіпофіз або на гіпоталамус за так званим від’ємним зворотним зв’язком. Підвищений рівень таких гормонів у кровоносному руслі приводить до зниження рівня в крові гормон-регулівних факторів і тропних гормонів гіпофіза і, навпаки, низький рівень у крові гормонів посилює вивільнення тропних гормонів.
Існують нервові зв’язки між гіпоталамусом і нейрогіпофізом, однак вони відсутні між гіпоталамусом та аденогіпофізом. Це пов’язано з тим, що гормони гіпоталамуса надходять в аденогіпофіз по портальних судинах (венулах), які розташовані в ніжці лійки. У разі порушення цього шляху доставка гормон-регулівних факторів з гіпоталамуса у гіпофіз припиняється.
Кровопостачання гіпоталамо-гіпофізної ділянки здійснюється гілками внутрішньої сонної артерії (верхньою та нижньою гіпофізними артеріями), а також гілками артеріального кола великого мозку. Верхні гіпофізні артерії кровопостачають аденогіпофіз, а нижні нейрогіпофіз, вступаючи у контакт з аксонами ядер гіпоталамуса. Верхні гіпофізні артерії входять у середній гіпоталамус, де розгалужуються, утворюючи капілярну сітку (первинне капілярне сплетення); в останнє всмоктуються гормон-регулівний фактор гіпоталамуса. Після контакту з терміналями аксонів великих нейросекреторних клітин ядер гіпоталамуса, ці капіляри збираються в портальні вени, які опускаються вздовж лійки гіпофіза і входять у паренхіму аденогіпофіза. Тут вони знову розділяються, утворюючи сітку синусоїдних венозних капілярів (вторинне капілярне сплетення).
Таким чином, кров потрапляє до аденогіпофіза, попередньо пройшовши через проміжне поле гіпоталамуса, де вона збагачується на гіпоталамічні аденогіпофізотропні гормони. Відтік крові, насиченої аденогіпофізними гормонами із численних капілярів вторинного капілярного сплетення, здійснюється по системі вен, що впадають у венозні пазухи твердої мозкової оболони (печеристі та міжпечеристі), а далі в загальний кровотік. Таким чином, ворітна система гіпоталамуса є морфофункціональним компонентом складного механізму нейрогуморального контролю тропних функцій аденогіпофіза.
Функції гіпоталамуса. Центральна нервова система рефлекторно здійснює контроль над автономним відділом нервової системи, який регулює діяльність внутрішніх органів через гіпоталамус; завдяки сигналам від контрольних систем гіпоталамуса, які надходять в центральні передвузлові автономні відділи провідних шляхів головного та спинного мозку. Також гіпоталамус регулює функції життєзабезпечення: обмін речовин і травлення, температуру тіла, водно-сольовий балансу, тиск і об’єм крові, сон, емоційну і статеву сфери:
i регуляція температури тіла здійснюється через рецептори шкіри, травної і дихальної систем, що призводить до стимуляції центрів гіпоталамуса, які посилюють обмін речовин;
i водно-сольовий баланс регулюється осморецепторами, гіпоталамусом за рахунок контролю тиску крові за участю гіпофіза, нирок і над надниркових залоз;
i центр насичення та голоду розташований у вентральному ядрі гіпоталамуса, імпульси надходять від механорецепторів травного шляху і глюкозорецепторів судин;
i тиск і об’єм крові регулюється гіпоталамусом через хемо- і барорецептори рефлексогенних зон аорти та порожнистих і яремних вен;
i у гіпоталамусі розташовані центри емоційної і статевої сфер; при подразненні вентромедіальних ядер у тварин виникає агресивна поведінка лють, немотивовані реакції;
i у підталамічній ділянці, на думку І.П. Павлова, розташований центр сну, проте, сучасними дослідженнями доведено, що сон є особливим станом клітин кори великого мозку, а зорові імпульси збуджують центри гіпоталамуса і сприяють пробудженню людини від сну;
i гіпоталамус забезпечує постійність внутрішнього середовища організму, тобто постійність осмотичного, гідростатичного і температурного станів.
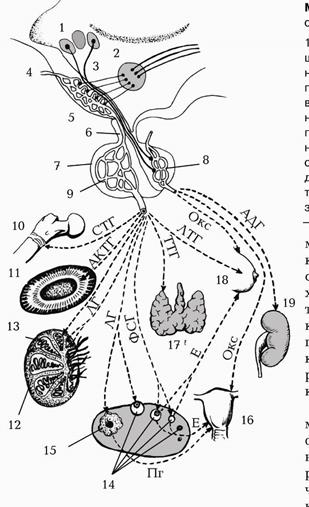
Мал. 226. Схема впливу гіпоталамогіпофізарної системи на органи.
1. Супраоптичне ядро гіпоталамуса. 2. Паравентрикулярне ядро гіпоталамуса. 3. Відростки нервових клітин ядер гіпоталамуса. 4. 5. 6. Аденогіпофіз. 7. Нейрогіпофіз. 8. 9. Метафізи кісток. 10. Кора наднирника. 11. Інтерстицій яєчка. 12. Сперматогенний епітелій яєчка. 13. Стадії дозрівання яйцеклітини в яєчнику. 14. Жовте тіло яєчника. 15. Матка. 16. Щитоподібна залоза. 17. Молочна залоза. 18. Нирка.
Дата добавления: 2017-01-13; просмотров: 1247;