Межуточный обмен липидов. Липогенез и липолиз в жировой ткани, их регуляция. Роль ЛП-, и ТАГ-липаз.
Процесс образования, отложения и мобилизации из депо жира регулируется нервной и эндокринной системами, а также тканевыми механизмами и тесно связаны с углеводным обменом. Так, повышение концентрации глюкозы в крови уменьшает распад триглицеридов и активизирует их синтез. Понижение концентрации глюкозы в крови, наоборот, тормозит синтез триглицеридов и усиливает их расщепление. Таким образом, взаимосвязь жирового и углеводного обменов направлена на обеспечение энергетических потребностей организма. При избытке углеводов в пище триглицериды депонируются в жировой ткани, при нехватке углеводов происходит расщепление триглицеридов с образованием неэстерифицнрованных жирных кислот, служащих источником энергии.Ряд гормонов оказывает выраженное влияние на жировой обмен. Сильным жиромобилизирующим действием обладают гормоны мозгового слоя надпочечников — адреналин и норадреналин, поэтому длительная адреналинемия сопровождается уменьшением жирового депо. Соматотропный гормон гипофиза также обладает жиромобилизирующим действием. Аналогично действует тироксин — гормон щитовидной железы, поэтому гиперфункция щитовидной железы сопровождается похуданием.Наоборот, тормозят мобилизацию жира глюкокортикоиды — гормоны коркового слоя надпочечника, вероятно, вследствие того, что они несколько повышают уровень глюкозы в крови.Обмен жировой ткани - липолиз и липогинез. Два процесса способствуют и противодействуют развитию ожирения: липогенез и липолиз. К регуляторам липогенеза относятся инсулин, простагландины, вазопрессин; к регуляторам липолиза - СТГ, АКТГ, катехоламины, половые гормоны и липотропины. Липогенез. Главными побуждающими факторами жирообразования служат поступающие в организм с пищей так называемые липогенные вещества, которыми в первую очередь являются простые углеводы (сахар, сладости, картофель, мучные и макаронные изделия) и только во вторую очередь, как это не странно, собственно сами животные и растительные жиры. На пищеварительное усвоение углеводов в сравнении с жирами требуется значительно меньше энергии и времени, поступивший в пищу сахар способен уже в течение нескольких десятков минут превратиться в жировые отложения на нашем теле. Это связанно с тем, что жиры и углеводы состоят из одних и тех же химических элементов, что в свою очередь позволяет простым углеводам чрезвычайно быстро модифицироваться в жирные кислоты. Сами жиры усваиваются значительно дольше, да и их усвоение связанно с потерей большего количества энергии, которая преимуществено получается от расщепления этих же липогенных веществ. Липолиз. Окончательное расщепление или окисление жиров происходит в клетках скелетной и сердечной мускулатуры, а также мозга и внутренних органов. В тех случаях, когда расщепляемые жирные кислоты поступают к местам их окончательного сжигания из жировой ткани, они должны быть сначала первично расщеплены в клетках их запасания и перенесены кровью к местам их непосредственного окисления. Этот процесс первичного расщепления или липидной мобилизации достаточно сложен и длителен, кроме того, активная мобилизация жира возможна только в жировых областях, обильно снабженных кровеносными капиллярами, те же места, где капиллярное русло развито менее значительно, вовлекаются в процесс мобилизации много позднее. Несколько ускоряют эти процессы физические нагрузки и тонизирующий массаж "проблемных" областей.
46. Биосинтез триацилглицеролов. Синтез жиров из углеводов. Регуляция процессов.Кроме пищевых жиров источником жирных кислот в организме служит их синтез из глюкозы. Непосредственным предшественником жирных кислот при их синтезе в организме является ацетил-КоА, т. е. то же вещество, которое образуется при р-окислении жирных кислот. Несмотря на то, что все реакции р-окисления обратимы, они не используются для синтеза жирных кислот.Ацетил-КоА для синтеза жирных кислот образуется путем окислительного де- карбоксилирования пирувата. Кроме того, окисление и синтез жирных кислот разделены в пространстве: окисление происходит в митохондриях, а синтез — в цитозоле.
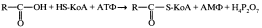
Большинство жирных кислот, необходимых человеку, может синтезироваться в организме из углеводов. Часть углеводов, поступающих с пищей, превращается в организме в жиры, особенно если количество углеводов превышает необходимое для возобновления запасов гликогена в печени и мышцах. Схема этого превращения представлена на рис. 10.15.
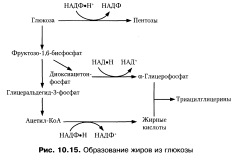
Глюкоза служит источником ацетил-КоА, из которого синтезируются жирные кислоты. Необходимый для восстановительных реакций НАДФН поставляется за счет окисления глюкозы в пентозофосфатном пути, а также за счет дегидрирования яблочной кислоты НАДФ-зависимой малатдегидрогеназой. Глице- рол-3-фосфат получается путем восстановления диоксиацетонфосфата — промежуточного продукта гликолиза Таким образом, из глюкозы образуется все, что необходимо для синтеза жиров.Синтез жиров из углеводов наиболее активно происходит в печени, жировой ткани и лактирующих молочных железах. Синтез триацилглицеринов из глицерол-3-фосфата и ацил-КоА представлен на схеме:

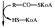
Лизофосфатидная кислота
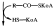
Фосфатидная кислота
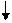 +Вода - Н3РО4
+Вода - Н3РО4
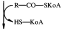 Диацилглицерин
Диацилглицерин
Триацилглицерин
47.Окисление жирных кислот. Значение, сущность, последовательность реакций β-окисления ВЖК. Энергетика процессов. Регуляция. Связь с ЦТК и ЦПЭ.
Специфический путь катаболизма жирных кислот (р-окисление). При р-окислении окисляется группа —СН2— в в-положении жирной кислоты до группы
 (рис. 10.4, реакции 1-3). При этом на двух стадиях происходит дегидрирование: при участии ацилдегидрогеназы (реакция 1, флавиновый фермент, водород переносится на убихинон) и (3-оксиацилдегидрогеназы (реакция 3, акцептор водорода НАД+). Затем р-кетоацил-КоА при действии фермента тиолазы (реакция 4) распадается на ацетил-КоА и ацил-КоА, укороченный на два углеродных атома по сравнению с исходным. Этот ацил-КоА вновь подвергается в-окислению.
(рис. 10.4, реакции 1-3). При этом на двух стадиях происходит дегидрирование: при участии ацилдегидрогеназы (реакция 1, флавиновый фермент, водород переносится на убихинон) и (3-оксиацилдегидрогеназы (реакция 3, акцептор водорода НАД+). Затем р-кетоацил-КоА при действии фермента тиолазы (реакция 4) распадается на ацетил-КоА и ацил-КоА, укороченный на два углеродных атома по сравнению с исходным. Этот ацил-КоА вновь подвергается в-окислению.
Многократное повторение этого процесса приводит к полному распаду жирной кислоты до ацетил-КоА.
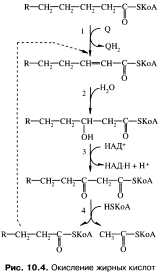
Например, молекула пальмитиновой кислоты (паль- митил-КоА), содержащая 16 углеродных атомов, превращается в 8 молекул ацетил- КоА за 7 циклов р-окисления. Образующиеся в реакциях дегидрирования восстановленные коферменты передают водород в дыхательную цепь: за счет этого при р-окислении 1 моль пальмитиновой кислоты может синтезироваться 35 моль АТФ. Использование жирных кислот путем р-окисления происходит во многих тканях. Особенно значительна роль этого источника энергии в скелетных мышцах при длительной физической работе и в сердечной мышце. Около 70 % кислорода, поглощаемого сердечной мышцей, используется для окисления жирных кислот. Нервная ткань не использует жирные кислоты как источник энергии.
В организме преобладают жирные кислоты с четным числом углеродных атомов. Из жирных кислот с нечетным числом углеродных атомов, имеющихся в организме в небольшом количестве, на завершающей стадии р-окисления образуется пропионил-КоА. Кроме того, пропионил-КоА образуется при распаде некоторых аминокислот (валина, изолейцина, треонина, метионина).Метаболизм всех липидов тесно связаны друг с другом
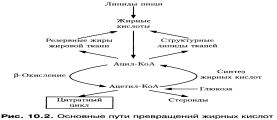
49.Биосинтез высших жирных кислот: последовательность реакций, локализация процесса, характеристика ферментов, регуляция.В органах и тканях человека синтезируются почти все необходимые жирные кислоты за исключением полиеновых (вжк). Исходным соединением для синтеза (вжк) является ацетилКоА, который в клетках может образовываться из различных соединений. Преимущественно используется ацетилКоА,образующийся при окислении моносахаридов.Одновременно используется ацетилКоА, который образуется при распаде углеродных скелетов аминокислот. Синтез (вжк) может протекать в клетках, однако основная масса соединений этого класса синтезируется в печени и жировой ткани. Важнейшим субстратом, продукты метаболизма которого используются для синтеза липидов, является глюкоза АцетилКоА используемый при липогенезе образуется в основном в матрикее митохондрий. Синтез высших жирных кислот идет в цитозоле Внутренняя мембрана митохондрий непроницаема для ацетилКоА поэтому существует система (челночная) транспорта ацетильных остатков из матрикса митохондрий в цитозоль. Синтез фосфолипидов. Все необходимые организму фосфолипиды могут синтезироваться в его клетках, причем в клетках могут функционировать несколько различных метаболических путей биосинтеза глицерофосфолипидов. Синтез лицитина А) при реакции ацилирования образуется фосфотидная кислота далее за счет фермента фосфотазы фосфотиднойкислоты образуется диглнцерид Б) Параллельно идет активация аминоспиртов (при наличии свободных аминоспиртов в клетке) далее идетактивация холина с образованием его производного - ЦДФхоли (активированный) он может включаться всинтезх.в) Диглицерид+ЦДФхолин - трансферазная реакция обеспечивает образование фосфотидилхолина и отщепляется ЦМФ, который затем за счет энергии АТФ может превращаться в ЦДФ. Алътернатнвным вариантоу синтеза может быть синтез с промежуточным образованием фосфотидной кислоты, но уже активированной. Сама фосфотидная кислота образуется при помощи ацилирования фосфоглицерина. Далее она взаимодействует с цитидин-3-фосфатом с образованием активной фосфотиднои кислоты (ЦДФ-фосфотидная кислота). Далее идет превращение в фосфотидилсерин или в инозитолфосфатид В том и другом случае происходит отщепление ЦМФ
Дата добавления: 2016-12-08; просмотров: 8066;