СТРУКТУРЫ МОЗГА, РЕАЛИЗУЮЩИЕ ПОДКРЕПЛЯЮЩУЮ, ПЕРЕКЛЮЧАЮЩУЮ, КОМПЕНСАТОРНО-ЗАМЕЩАЮЩУЮ И КОММУНИКАТИВНУЮ ФУНКЦИИ ЭМОЦИЙ
Результаты нейрофизиологических экспериментов показывают, что потребности, мотивации и эмоции имеют различный морфологический субстрат. Так, при стимуляции зон самораздражения латерального гипоталамуса градуально нарастающим электрическим током поведенческие реакции крысы всегда имеют одну и ту же последовательность. Слабое раздражение вызывает генерализованное поисковое поведение без обращения к находящимся в камере целевым объектам – пище, воде, педали для самораздражения. При усилении интенсивности тока животное начинает есть, пить, грызть и т.п. По мере дальнейшего усиления тока возникает реакция самораздражения с сопутствующими мотивационными эффектами, а затем – только самораздражение. Введением определенных фармакологических веществ в зоны самораздражения можно избирательно повлиять на мотивационные и эмоциональные компоненты поведения, например ослабить стремление животного к пище, не изменив объем ее поглощения в случае контакта [Ikemoto, Panksepp, 1996].
В зонах самораздражения латеральной преоптической области и латерального гипоталамуса зарегистрирована активность нейронов, специфически изменяющих свою активность при мотивационных и эмоционально позитивных состояниях, вызываемых электрическими и натуральными стимулами, изменением уровня пищевой и водной мотивации. Нейроны первого типа (мотивационные) обнаружили максимум активации при силе тока, вызывающей мотивационное поведение, и тормозили свою активность при снижении уровня естественной мотивации. Нейроны второго типа (подкрепляющие) достигали максимума активации при стимуляции силой тока, вызывающей самораздражение, и активировались при насыщении. Мотивационные и эмоционально позитивные состояния находятся между собой в конкурентных отношениях и вызывают противофазные перестройки нейронов первого и второго типов [Зайченко и др., 1995].
Как показали эксперименты Н.Г. Михайловой и М.И. Зайченко в лаборатории П.В. Симонова, нейроны этих двух типов по-разному участвуют в реализации классических и инструментальных оборонительных условных рефлексов у крыс, когда условный сигнал (свет) сочетается с эмоционально негативным внутримозговым раздражением дорзолатеральной покрышки. На рис. 7.1 представлены изменения импульсной активности мотивационных (наверху) и позитивно подкрепляющих (внизу) нейронов латерального гипоталамуса при осуществлении условных оборонительных рефлексов: классического, избавления и избегания. Столбики последовательно показывают частоту разрядов за 5 с до условного стимула, при действии света, при действии света совместно с током и после выключения стимулов. Как следует из рисунка, осуществление классического условного рефлекса и реакции избавления сопровождается усилением активности мотивационных нейронов и подавлением импульсации подкрепляющих. Только хорошо выработанная реакция избегания, при которой крыса перестает получать наказание электрическим током, ведет к усилению активности позитивно подкрепляющих нейронов. Эти данные позволяют ответить на давно обсуждаемый в литературе вопрос: что именно служит подкреплением инструментальных оборонительных рефлексов? В случае классических рефлексов и реакций избавления подкреплением служит эмоционально негативное состояние страха. При успешном осуществлении реакции избегания в процесс вовлекается механизм положительных эмоций.
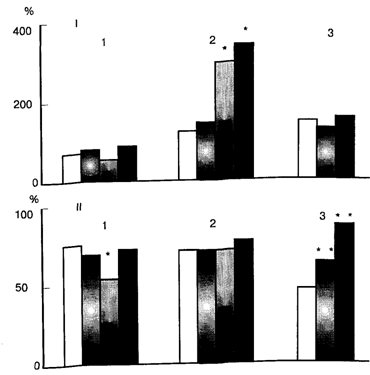
Рис.7.1 . Изменения импульсной активности нейронов латерального гипоталамуса крыс при осуществлении оборонительных условных рефлексов - классического (1) избавления (2) и избегания (3). По вертикали - средняя частота разрядов в процентах к фону. I - мотивационная. II - позитивно подкрепляющая группа нейронов. Столбики последовательно представляют частоту разрядов за 5 с до включения стимулов, при действии условного света, при действии света и тока, после выключения стимулов. Одна звездочка - р меньше 0,05; две звездочки - р меньше 0,01
Оборонительные реакции нападения или бегства приурочены к различным структурам гипоталамуса. Стимуляция пунктов медиального гипоталамуса вызывающая оборонительные реакции кролика, ведет к возрастанию количества пар скоррелированно работающих нейронов зрительной и сенсомоторной коры, причем опережение разрядами нейронов последней разрядов нейронов зрительной коры достигает 120 мс. Этот факт можно рассматривать как своеобразную подготовку зрительной области к восприятию значимых для оборонительного поведения сигналов по механизму обратных связей Э.А. Асратяна или реинтеринга Дж.Эделмена [Павлова и др., 1995].
Вовлечение эмоционально-позитивных структур в реализацию реакций избегания (но не избавления!) позволяет понять, почему сам факт активного сопротивления неблагоприятно сложившейся ситуации (например, возможность для фиксированной на спине крысы кусать палочку) достоверно ослабляет объективно регистрируемые проявления стресса.
Ранее уже говорилось о том, что влияние эмоций на поведение определяется отношением животного к своему эмоциональному состоянию, причем подчинено принципу максимизации положительных эмоций и минимизации отрицательных. Этот принцип реализуется влиянием мотивационно-эмоциональных структур гипоталамуса на информационные (когнитивные) и организующие движение отделы новой коры, о чем свидетельствует анализ пространственной синхронизации электрической активности мозговых структур при самостимуляции крыс слабым постоянным током [Pavlygina et al., 1976].
На рис 7.2 отображен процент случаев регистрации значимой когерентности (Р < 0,05) на частотах альфаи тета-диапазонов в потенциалах сравниваемых мозговых образований. Рассматривая рисунок, можно убедиться, что непосредственно перед нажатием на педаль резко (более чем в 3 раза) увеличивается процент случаев статистически значимой когерентности электрической активности эмоционально позитивного пункта гипоталамуса с моторной областью коры. Возросшая когерентность свидетельствует о готовности нервных путей к проведению возбуждения по трем каналам условных связей: от ранее раздражавшегося эмоционально позитивного пункта к двигательной коре, к зрительной коре, а также между зрительным анализатором, воспринимающим условный сигнал будущего подкрепления (вид педали, ее местонахождение в камере и т.п.), и двигательной областью коры, поскольку именно вид педали будет направлять движение животного, инициированное следовым возбуждением эмоционально позитивной зоны.
В период пребывания крысы на педали когерентность уменьшается, животное получает подкрепление и становится совершенно пассивным. Непосредственно перед уходом впервые увеличивается когерентность негативного пункта и моторной области коры: возбуждение негативных структур готово к трансформации в двигательную реакцию избавления. После ухода животного с педали процент случаев достоверного увеличения когерентности возвращается к исходным цифрам. Только следы эмоционально негативного состояния все еще обнаруживаются при сопоставлении электрической активности негативного пункта с двигательной областью коры.
Как показали исследования Р.А. Павлыгиной и Ю.В. Любимовой [1994], мотивационные влияния гипоталамуса на новую кору носят асимметричный характер. Эта асимметрия проявляется после односуточной пищевой депривации кроликов не только в преимущественной активации левого полушария, но и в усилении взаимосвязи электрических процессов левого полушария. Анализ спектрально-корреляционных характеристик электрической активности мозга кролика обнаружил большую реактивность левого полушария по сравнению с правым и при состоянии жажды [Маликова, Мац, 1991]. Преобладающая активность левого полушария при возникновении голода у человека зарегистрирована в альфаи дельта-диапазонах его электроэнцефалограммы [Сулимов, 1995].
О межполушарной асимметрии при естественной пищевой мотивации свидетельствует регистрация импульсной активности отдельных нейронов зрительной и сенсомоторной области неокортекса кроликов, а также взаимодействие этих нейронов. Судя по импульсации нейронов, кора левого полушария более активна у голодных кроликов, а кора правого полушария – у сытых. Наиболее выраженные различия наблюдались в активности нейронов фронтальных областей, наименее выраженные – в сенсомоторных нейронах [Павлова, 1995]. В ходе этих исследований была впервые установлена функциональная асимметрия самого «пищевого» гипоталамуса. При стимуляции левого гипоталамуса пороги пищевых реакций были меньше, а сами реакции более стабильны, чем при стимуляции правого гипоталамуса, причем «пищевая» асимметрия латерального гипоталамуса не коррелировала с моторной и сенсорной асимметрией у этих животных [Павлова, Мац, 1996].
Вся совокупность имеющихся в настоящее время данных свидетельствует о том, что гипоталамус является ключевой структурой для реализации наиболее древней подкрепляющей функции эмоций, для решения универсальной поведенческой задачи максимизации-минимизации возникшего эмоционального состояния: приближения или избегания. Ведь именно получение желательных, эмоционально положительных стимулов или устранение нежелательных, эмоционально отрицательных, а не удовлетворение какой-либо потребности служит непосредственным подкреплением при обучении. У крыс невозможно выработать инструментальный условный рефлекс при введении пищи через канюлю в желудок (т.е. минуя вкусовые рецепторы), хотя такой рефлекс вырабатывается при введении в желудок морфина, который очень быстро вызывает у животного положительное эмоциональное состояние. Тот же морфин из-за своего горького вкуса перестает быть подкреплением, если его вводить через рот [Cytawa, Trojniar, 1976]. В другой серии опытов авторы вырабатывали инструментальный пищевой условный рефлекс у крыс, а после его упрочения заменяли натуральную пищу введением питательного раствора в желудок через носоглоточную канюлю. Рефлекс нажатия на рычаг при этом угасал, но сохранялся, если в желудок вводили 0,05%-ный раствор морфина [Trojniar, Cytawa, 1976].
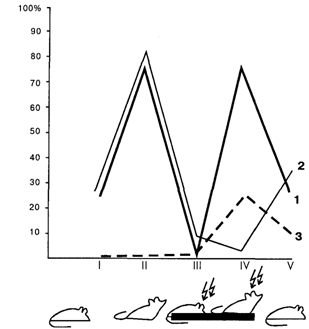
Рис. 7.2. Процент случаев статистически значимого изменения когерентности электрической активности мозговых структур:
/– спокойное состояние; //– перед посадкой на педаль; ///– на педали; IV– перед уходом с педали; V – сразу же после ухода; 1 – эмоционально положительный пункт гипоталамуса – двигательный неокортекс; 2– эмоционально отрицательный пункт гипоталамуса – двигательный неокортекс; 3 – двигательная область – зрительная область неокортекса
В отличие от гипоталамуса, сохранность второго «мотивационного» мозгового образования – миндалины – необязательна для формирования сравнительно простых форм временных связей. По данным М.Л. Пигаревой [1983], двухсторонняя амигдалоэктомия не препятствует выработке пищевых и оборонительных условных рефлексов. При этом миндалина «безразлична» к вероятности подкрепления, к его информационному компоненту. Скорость выработки условных двигательных пищевых рефлексов у амигдалэктомированных крыс не отличается от аналогичных показателей у интактных животных ни при высокой (100 и 50%), ни при низкой (25%) вероятности их подкрепления. Только вероятность в 33%, для которой характерен максимум возникающего при этом эмоционального напряжения, сопровождается некоторым замедлением процесса формирования пищедобывательной реакции. Разрушение миндалин у крыс через неделю после однократного обучения реакции избегания не препятствует воспроизведению этого навыка через четыре дня после операции [Parent et al., 1995]. При одностороннем выключении правой или левой миндалины через 24 ч или 10 дней после сеанса выработки реакции избегания оказывается, что время задержки реакции короче в том случае, если была выключена правая миндалина [Coleman-Mesches, McGaugh, 1995].
Результаты опытов на животных совпадают с клиническими наблюдениями. У двух амигдалэктомированных пациентов не отмечено нарушений формирования ассоциаций между объектами двух разных сенсорных модальностей [Lee et al., 1995]. Двухстороннее поражение миндалин не приводит к амнезии. Дефект памяти носит строго избирательный характер, связанный с эмоциональной окраской событий [Markowitsch et al., 1994].
Сохранность миндалины наиболее существенна в том случае, когда возникает конкуренция между сосуществующими мотивациями и становится необходимым выбор доминирующей потребности, подлежащей удовлетворению в данный момент. Хорошей экспериментальной моделью подобной ситуации может служить выработка условно-рефлекторного переключения разнородных условных рефлексов по Э.А. Асратяну [Пигарева, 1983], когда один и тот же сигнал (звук) утром подкрепляется пищей, а вечером – болевым раздражением. Судя по проценту правильных оборонительных и пищевых условных реакций, на протяжении 40 дней не удалось выработать условнорефлекторное переключение у амигдалэктомированных крыс. Вместе с тем решение подобной поведенческой задачи возможно, если искусственно создать достаточный дисбаланс между конкурирующими мотивациями и соответствующими им эмоциями: между голодом и страхом (табл.).
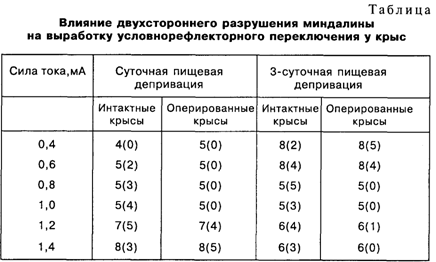
Цифры без скобок показывают количество крыс в каждой группе, а цифры в скобках – то количество животных, у которых удалось выработать условно-рефлекторное переключение (три опыта подряд – со 100-процентным выполнением как пищевых, так и оборонительных условных рефлексов) на протяжении 60 опытных дней. Амигдалэктомированные крысы справляются с этой задачей, если сильное болевое раздражение сочетается с односуточной пищевой депривацией или, напротив, слабое болевое раздражение применяется при сильном голоде после трехсуточной депривации. Иными словами, миндалина играет решающую роль в осуществлении переключающей поведение функции эмоций, т.е. в выборе мотивации, которая соответствует не только той или иной потребности, но и внешним условиям ее удовлетворения в данной ситуации и в данный момент.
Судя по новейшим данным, полученным с помощью позитронно-эмиссионной томографии, миндалина реализует свою переключающую функцию через хвостатое ядро. Миндалина вовлекается в процесс организации поведения на сравнительно поздних его этапах, когда актуализированные потребности уже сопоставлены с перспективой их удовлетворения и трансформированы в соответствующие эмоции. Так, введение никотина в миндалину крыс не влияло на потребление воды и соли при свободном доступе к воде и пище. У лишенных воды животных инъекция никотина в базолатеральное ядро миндалины блокировала потребление соли [Saad et al., 1995]. Поскольку неофобия представляет конкуренцию между пищевой и ориентировочно-оборонительной мотивацией и определяется потоками информации из миндалины и гиппокампа в прилежащее ядро, становится понятно, почему разрушение миндалины ослабляет страх перед незнакомой пищей [Burns et al., 1996].
Что касается прогнозирования вероятности удовлетворения потребности (вероятности подкрепления), оно реализуется «информационными» структурами мозга – гиппокампом и фронтальными отделами новой коры.
Наиболее ярким дефектом гиппокампэктомированных животных оказалась их чувствительность к ситуациям с низкой вероятностью подкрепления условных сигналов (рис. 7.3). При вероятности подкрепления пищевых условных рефлексов, равной 100 и 50%, гиппо-кампэктомированные крысы хотя и отстают от интактных, но все же справляются с задачей. Выработка условных рефлексов при вероятности подкрепления 33 и 25% оказалась для них недоступной. Напомним, что в опытах с условно-рефлекторным переключением вероятность подкрепления звука пищей является высокой в утренних опытах и низкой – в вечерних, а вероятность подкрепления того же звука болевым раздражением носит прямо противоположный характер. После десяти дней безуспешных попыток выработать переключение оборонительных и пищевых условных рефлексов у крыс двухсторонняя гиппокампэктомия привела к формированию стабильного условно-рефлекторного переключения. Двухсторонняя гиппокампэктомия не только облегчает выработку условно-рефлекторного переключения, но и устраняет признаки эмоционального напряжения у этих животных, о чем судили по изменениям частоты сердцебиений. Повреждение дорзального гиппокампа крыс сказывается на обстановочном оборонительном условном рефлексе, не влияя на высоковероятную связь звукового стимула с болевым раздражением лап.
Способность гиппокампа реагировать на сигналы маловероятных событий позволяет рассматривать его как ключевую структуру для реализации компенсаторной (замещающей недостаток информации) функции эмоций. Эта функция проявляется не только в гипермобилизации вегетативных сдвигов (учащение сердцебиений, подъем кровяного давления, выброс в кровяное русло гормонов и т.п.), как правило, превышающих реальные нужды организма. Возникновение эмоционального напряжения сопровождается переходом к иным, чем в спокойном состоянии, формам поведения, механизмам оценки внешних сигналов и реагирования на них в соответствии с принципом доминанты А.А. Ухтомского. Не случайно ученик И.П. Павлова психиатр В. П. Осипов назвал «эмоциональной» первую стадию выработки условного рефлекса – стадию генерализации, которая по своим поведенческим, электрофизиологическим и нейроанатомическим характеристикам совпадает с проявлениями доминанты А.А. Ухтомского [Pavlygina, 1991, 1991а]. Эмоционально возбужденный мозг реагирует на широкий круг предположительно значимых сигналов, истинное значение которых – соответствие или несоответствие реальной действительности – выясняется лишь позднее по мере стабилизации условного рефлекса.
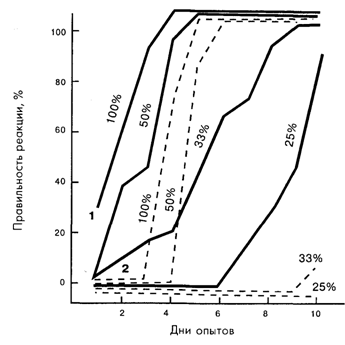
Рис. 7.3. Процент правильных условных пищевых реакций у контрольных (1) и гиппокампэктомированных (2) крыс при различной вероятности подкрепления условного сигнала
Если процесс упрочения условного рефлекса сопровождается уменьшением эмоционального напряжения и одновременно переходом от доминантного (генерализованного) реагирования к строго избирательным реакциям на условный сигнал, то возникновение эмоций ведет к вторичной генерализации. Нарастание эмоционального напряжения, с одной стороны, расширяет диапазон извлекаемых из памяти энграмм, а с другой – снижает критерии «принятия решения» при сопоставлении этих энграмм с наличными стимулами. Чем сильнее тревога, тем чаще субъект отвечает на нейтральный стимул как на аверсивный.
Предположительное доминантное реагирование целесообразно только в условиях прагматической неопределенности. Недостающая информация пополняется путем поискового поведения, совершенствования навыков, мобилизации хранящихся в памяти энграмм. Компенсаторное значение отрицательных эмоций заключается в их замещающей роли. Что касается положительных эмоций, то их компенсаторная функция реализуется через влияние на потребность, инициирующую поведение. В трудной ситуации с низкой вероятностью достижения цели даже небольшой успех (увеличение вероятности) порождает положительную эмоцию, которая усиливает потребность согласно правилу, вытекающему из «формулы эмоций».
В отличие от гиппокампа, вторая «информационная» структура мозга – фронтальная новая кора – ориентирует поведение на сигналы высоковероятных событий. На рис. 7.4А показан процент двигательных пищевых условных реакций у интактных крыс при различной вероятности подкрепления. Видно, что выработка условного рефлекса происходит тем медленнее, чем ниже вероятность подкрепления. После разрушения передних отделов новой коры (рис. 7.4Б) процесс формирования условного рефлекса при низкой вероятности подкрепления заметно ускоряется. Иными словами, сигналы с различной вероятностью их подкрепления пищей становятся одинаково эффективными. Этот результат представляет тем больший интерес, что, судя по имеющимся данным, фронтальные области коры головного мозга крыс не отличаются по своим основным функциям от фронтальной коры высших позвоночных, включая приматов.
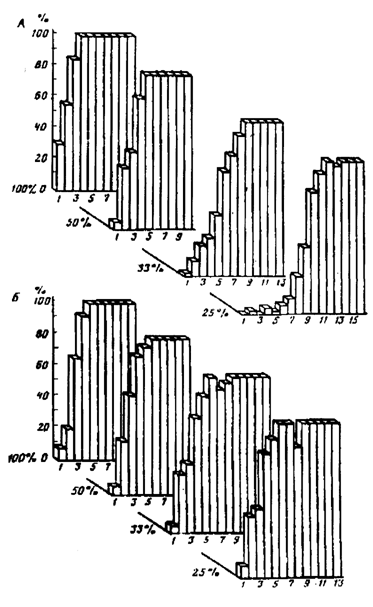
Рис. 7.4. Процент двигательных пищевых условных реакций у контрольных (А) и лобэктомированных (Б) крыс при различной вероятности подкрепления (по оси абцисс – дни опытов)
Отметим, что у собак нарушение вероятностного прогнозирования наблюдается после удаления дорсолатеральных, а не медиобазальных областей префронтальной коры. Такому разделению функций соответствуют анатомические связи этих двух отделов фронтальной коры: дорсального – с гиппокампом, а вентрального – с миндалиной. Префронтальная кора обезьян получает зрительную информацию из нижневисочной коры, слуховую – из верхней, а соматосенсорную – из задней теменной. Информация, связанная с мотивацией и памятью, поступает из гипоталамуса, миндалины и гиппокампа. Префронтальная кора имеет две функции: извлечение из памяти необходимой информации и организация целенаправленного поведения путем взаимодействия с премоторной корой и базальными ганглиями. Блокада переднего стриатума и переднего бледного шара нарушает приобретение нового навыка у обезьян при сохранении ранее выработанного. Блокада заднего стриатума нарушает выполнение выработанного навыка, не препятствуя приобретению нового.
В префронтальной коре приматов обнаружены нейроны, связанные с прогнозированием местоположения пищи и ее качества. Так, активность нейронов префронтальной коры увеличивается сильнее, если ожидается предпочитаемая пища. Регистрация активности различных отделов префронтальной коры у человека с помощью позитронно-эмиссионной томографии показала, что при мысленном представлении эмоционально нейтральных событий преимущественно активируются дорсолатеральная префронтальная кора и задняя височная область, в то время как при мысленном представлении эмоционально окрашенных сцен более активны медиальная префронтальная кора и передняя височная область.
Интенсивность эмоционального напряжения безотносительно к его знаку связывают с активностью теменновисочных отделов правого полушария. Именно от нее зависит выход эмоционального напряжения на вегетативные функции, проявляясь сдвигами кожногальванического рефлекса, частоты сердечных сокращений, кровяного давления, секреции кортизона и т.п. Важная роль височной коры правого полушария в осуществлении эмоциональных реакций обнаружена и у животных. Что касается знака эмоций (положительного или отрицательного), то, по мнению Дейвидсон [Davidson, 1992] и Геллер [Heller, 1993], он зависит от соотношения активности левой фронтальной коры (ЛФК) и правой фронтальной коры (ПФК). Это правило В. Геллер представила в виде двух неравенств: ЛФК > ПФК – положительные эмоции; ПФК > ЛФК – отрицательные эмоции. При подаче субъекту неосознаваемых (невербализуемых) стимулов правое полушарие более чувствительно к эмоционально отрицательным, а левое – к эмоционально положительным.
В упомянутых работах остался без ответа вопрос о том, чем определяется специфика левой и правой фронтальной коры в генезе положительных и отрицательных эмоций? Было бы упрощением предполагать, что в этих двух мозговых образованиях непосредственно локализованы «центры» соответствующих эмоций. Согласно потребностно-информационной теории эмоций положительная эмоция возникает, когда имеющаяся информация превышает прогностически необходимую, а отрицательная – когда необходимая больше имеющейся. Если сопоставить эти неравенства с неравенствами В. Геллер, приведенными ранее, то можно сделать вывод о преимущественной связи ПФК с прагматической информацией, необходимой для удовлетворения потребности, т.е. приобретенной ранее и хранящейся в памяти, а ЛФК – с информацией, имеющейся в данный момент, которая поступила недавно. Это предположение имеет ряд экспериментальных доказательств.
Судя по электрической активности мозга, на первом этапе выработки классического условного рефлекса у собак преобладает левое полушарие, а по мере его закрепления – правое. Аналогичные данные получены и у человека с помощью позитронно-эмиссионной томографии. Ранее мы уже приводили данные о том, что возникновение мотивации сопровождается повышенной активностью передних отделов левого полушария. Вся совокупность полученных в настоящее время данных позволяет утверждать, что «закон Геодакяна» [Геодакян, 1992] о формировании в процессе эволюции новых функций первоначально в левом полушарии с последующим их перемещением в правое справедлив не только для филогенеза, но и для процесса индивидуального обучения. Именно учет специфики информационных (когнитивных) функций, осуществляемых левым и правым фронтальным неокортексом, позволяет ответить на вопрос о латерализации положительных и отрицательных эмоций и о той роли, которую эти мозговые структуры играют в генезе эмоциональных состояний.
Исключительно велика коммуникативная функция эмоций – порождение и восприятие выражения эмоций в мимике, голосе, физических характеристиках речи. По мнению ряда исследователей, около 90% эмоционального общения происходит на неречевом (несемантическом) уровне. При восприятии речи независимо от ее содержания мы можем судить об эмоциональном состоянии партнера (например, об аффектах тоски или тревоги) по таким показателям, как средняя длительность чистой речи, продолжительность пауз, отношение времени пауз к общему времени высказывания и скорость артикуляции [Фролов и др., 1994]. Подчеркнем, что оценка эмоционального состояния партнера по его мимике имеет собственный мозговой механизм, отличный от механизма идентификации партнера. Так двухстороннее повреждение височной коры нарушает узнавание знакомого лица, а одностороннее повреждение правой височной области достаточно для нарушения распознавания эмоциональной экспрессии. Двухстороннее повреждение миндалин, напротив, препятствует распознаванию мимики страха, не влияя на идентификацию знакомых и незнакомых лиц [Adolphs et al., 1995], а также интонационных признаков страха и гнева.
В процессах порождения и восприятия эмоциональной мимики мы вновь встречаемся с функциональной асимметрией мозга. Эмоциональное состояние субъекта преимущественно отражается на мимике левой половины лица, что свидетельствует о преобладающей активности правого полушария. Аналогичные данные получены и в опытах на макаках, что указывает на общность обнаруженной закономерности для всех приматов. Естественно, что эмоции распознаются лучше на фотографиях лиц, составленных из двух левых половин изображения. Правое полушарие (его центральная височная область) преобладает и при восприятии эмоциональной мимики. Правда, тщательно выполненное исследование обнаруживает достаточно сложную и динамичную картину взаимодействия полушарий при опознании эмоциональной лицевой экспрессии. При правильном опознании выражений радости, печали или эмоционально нейтральных лиц последовательно активируются фронтальная кора правого полушария, а затем – фронтальная кора левого. При ошибочных опознаниях активация левого полушария опережает активацию правого [Михайлова и др., 1996].
Завершая обзор взаимодействия передних отделов новой коры, гиппокампа, миндалины и гипоталамуса, мы приходим к выводу о том, что оно необходимо и достаточно для организации поведения в системе координат «потребности – вероятность их удовлетворения» с вовлечением механизмов, реализующих регуляторные функции эмоций. Что касается согласованной деятельности этих структур, то к ним в полной мере относится глубокая мысль А. А. Ухтомского о том, что «соподчинение физиологических приборов в порядке их нервной увязки есть процесс вынужденный и не предполагает для себя вмешательства какого-либо дополнительного, специального «координирующего центра» [Ухтомский, 1954].
На рис. 7.5 представлена схема взаимодействия мозговых структур в процессе организации поведенческого акта. Внутренние (метаболизм) и внешние (боль, запах и т.п.) побуждающие стимулы активируют мотивационные структуры гипоталамуса (ГТ), который, в свою очередь, активирует гиппокамп (ГИП) и передние отделы новой коры. Благодаря гиппокампу широкий круг внешних стимулов усиливает доминантное состояние. В случае совпадения этих стимулов с действием подкрепляющих факторов гиппокамп оказывается первым местом «встречи» сочетаемых афферентаций. При сформированном поведенческом акте в результате совместной деятельности гиппокампа и фронтальной коры (ФК) отбираются те внешние стимулы или их энграммы, которые ранее сопровождались удовлетворением данной потребности. Путем сопоставления мотивационного возбуждения с наличными стимулами и с энграммами, извлеченными из памяти, в миндалине (М) формируется эмоциональная окраска этих стимулов и энграмм, что ведет к выделению доминирующей мотивации, подлежащей первоочередному удовлетворению. Сложившаяся во фронтальной коре программа поступает в базальные ганглии, где путем взаимодействия с теменной корой вписывается в пространственные координаты предстоящего двигательного акта. Отсюда через моторную кору возбуждение поступает на эффекторные органы, реализующие целенаправленное поведение.
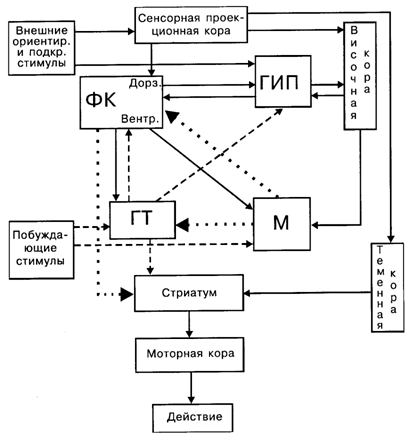
Рис. 7.5.Схема взаимодействия мозговых структур в процессе организации поведенческого акта: ФК– фронтальная кора; ГИП– гиппокомп; М– миндалина; ГТ– гипоталамус; сплошные линии – информационная афферентация, пунктирные – мотивационные влияния, точечные – эмоционально окрашенная афферентация
Исследование последствий хирургического, иммунологического или фармакологического выключения подробно описанных ранее мозговых образований привело к представлению о том, что индивидуальные особенности соотношения «информационной» системы (фронтальная кора и гиппокамп) с «мотивационной» системой (миндалина и гипоталамус) лежат в основе параметра экстра-интроверсии. Соотношение систем «фронтальная кора – гипоталамус» и «гиппокамп – миндалина» определяет другой параметр индивидуальных особенностей поведения, близкий по своим характеристикам параметру невротицизма – эмоциональной стабильности (рис. 7.6). Гипотеза о том, что отношения между так называемой информационной системой (неокортекс и гиппокамп) и мотивационной системой (миндалина и гипоталамус) представляют биологический субстрат интроверсии – экстраверсии и что отношения между новой корой и гипоталамусом, с одной стороны, и гиппокампом и миндалиной, с другой, формируют основу невротицизма, получила признание. Причем считается, что хотя эти идеи извлечены главным образом из опытов на крысах, они определенно обеспечивают достоверность концепции [Orebeke, 1988].
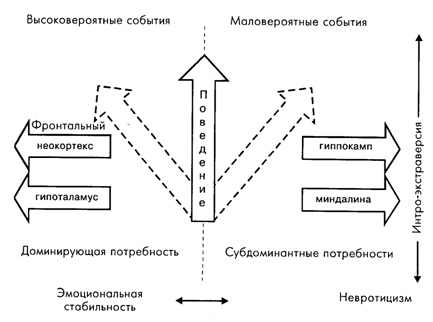
Рис. 7.6. Гипотетическая схема зависимости параметров интро-экстраверсии и эмоциональной стабильности – невротицизма от индивидуальных особенностей взаимодействия мозговых структур
Дата добавления: 2016-06-13; просмотров: 653;