НЕРВНАЯ СИСТЕМА РЫБ
Нервная система связывает организм с внешней средой и регулирует деятельность внутренних органов.

Рис. 1.Строение головного мозга (лосось)
А – вид сверху; Б – вид снизу; В – вид сбоку
1 – передний мозг; 2 – промежуточный мозг; 3 – средний мозг (зрительные доли); 4 – мозжечок; 5 – продолговатый мозг; 6 – обонятельные доли; 7 – эпифиз; 8 – гипофиз; 9 – сосудистый мешок
Нервная система представлена:
1) центральной (головной и спинной мозг);
2) периферической (нервы, отходящие от головного и спинного мозга).
Периферическая нервная система делится на:
1) соматическую (иннервирует поперечнополосатую мускулатуру, обеспечивает чувствительность тела, состоит из нервов, отходящих от спинного мозга);
2) вегетативную (иннервирует внутренние органы, делится на симпатическую и парасимпатическую, состоит из нервов, отходящих от головного и спинного мозга).
Головной мозг рыб включает пять отделов:
1) передний мозг (telencephalon);
2) промежуточный мозг (diencephalon);
3) средний мозг (mesencephalon);
4) мозжечок (cerebellum);
5) продолговатый мозг (myelencephalon) (рис. 1).
Внутри отделов головного мозга находятся полости.Полости переднего, промежуточного и продолговатого мозга называются желудочками, полость среднего мозга – сильвиевым водопроводом (она соединяет полости промежуточного и продолговатого мозга).
Передний мозг у рыб представлен двумя полушариями с неполной перегородкой между ними и одной полостью. В переднем мозге дно и бока состоят из нервного вещества, крыша у большинства рыб эпителиальная, у акул она состоит из нервного вещества. Передний мозг является центром обоняния, регулирует функции стайного поведения рыб. Выросты переднего мозга образуют обонятельные доли (у хрящевых рыб) и обонятельные луковицы (у костистых рыб).
В промежуточном мозге дно и боковые стенки состоят из нервного вещества, крыша – из тонкого слоя соединительной ткани. В нем различают три части:
1) эпиталамус (надбугровая часть);
2) таламус (средняя, или бугровая часть);
3) гипоталамус (подбугровая часть).
Эпиталамус образует крышу промежуточного мозга, в задней его части расположен эпифиз (железа внутренней секреции). У миног
здесь размещены пинеальный и парапинеальный органы, выполняющие светочувствительную функцию.
У рыб парапинеальный орган редуцируется, а пинеальный превращается в эпифиз.
Таламус представлен зрительными буграми, размеры которых связаны с остротой зрения. При слабом зрении они небольшие или отсутствуют.
Гипоталамус образует нижнюю часть промежуточного мозга и включает воронку (полый вырост), гипофиз (железа внутренней секреции) и сосудистый мешочек, где образуется жидкость, заполняющая желудочки головного мозга.
Промежуточный мозг служит первичным зрительным центром, от него отходят зрительные нервы, которые впереди воронки образуют хиазму (перекрест нервов). Также этот промежуточный мозг является центром переключения возбуждений, которые поступают из всех отделов мозга, связанных с ним, а через гормональную деятельность (эпифиз, гипофиз) участвует в регуляции метаболизма.
Средний мозг представлен массивным основанием и зрительными долями. Крыша его состоит из нервного вещества, имеет полость – сильвиев водопровод. Средний мозг является зрительным центром, также регулирует тонус мышц и равновесие тела. От среднего мозга отходят глазодвигательные нервы.
Мозжечок состоит из нервного вещества, отвечает за координацию движений, связанных с плаванием, сильно развит у быстроплавающих видов (акула, тунец). У миног мозжечок развит слабо и не выделяется в самостоятельный отдел. У хрящевых рыб мозжечок представляет полый вырост крыши продолговатого мозга, который сверху налегает на зрительные доли среднего мозга и на продолговатый мозг. У скатов поверхность мозжечка разделена бороздами на 4 части.
В продолговатом мозге дно и стенки состоят из нервного вещества, крыша образована тонкой эпителиальной пленкой, внутри его расположена полость желудочка. От продолговатого мозга отходит большинство головных нервов (с V по X), иннервирующих органы дыхания, равновесия и слуха, осязания, органы чувств системы боковой линии, сердце, пищеварительную систему. Задний отдел продолговатого мозга переходит в спинной мозг.
Рыбы в зависимости от образа жизни имеют различия в развитии отдельных отделов головного мозга. Так, у круглоротых хорошо развит передний мозг с обонятельными долями, слабо развит средний мозг и недоразвит мозжечок; у акул – хорошо развит передний мозг, мозжечок и продолговатый мозг; у костистых пелагических подвижных рыб с хорошим зрением – наиболее развиты средний мозг и мозжечок (скумбрия, летучая рыба, лососи) и т.д.
У рыб от головного мозга отходит 10 пар нервов:
I. Обонятельный нерв (nervus olfactorius) отходит от переднего мозга. У хрящевых и некоторых костистых обонятельные луковицы примыкают непосредственно к обонятельным капсулам и соединяются с передним мозгом при помощи нервного тракта. У большинства костистых рыб обонятельные луковицы примыкают к переднему мозгу, а от них к обонятельным капсулам идет нерв (щука, окунь).
II. Зрительный нерв (n. opticus) отходит от дна промежуточного мозга и образует хиазму (перекрест), иннервирует сетчатку глаза.
III. Глазодвигательный нерв (n. oculomotorius) отходит от дна среднего мозга, иннервирует одну из глазных мышц.
IV. Блоковый нерв (n. trochlearis) начинается от крыши среднего мозга, иннервирует одну из глазных мышц.
Все остальные нервы начинаются от продолговатого мозга.
V. Тройничный нерв (n. trigeminus) разделяется на три ветви, иннервирует челюстную мускулатуру, кожу верхней части головы, слизистую ротовой полости.
VI. Отводящий нерв (n. abducens) иннервирует одну из глазных мышц.
VII. Лицевой нерв (n. facialis) имеет много ветвей, иннервирует отдельные части головы.
VIII. Слуховой нерв (n. acusticus) иннервирует внутреннее ухо.
IX. Языкоглоточный нерв (n. glossopharyngeus) иннервирует слизистую оболочку глотки, мускулатуру первой жаберной дуги.
X. Блуждающий нерв (n. vagus) имеет много ветвей, иннервирует мускулатуру жабр, внутренние органы, боковую линию.
Спинной мозг расположен в спинномозговом канале, образованном верхними дугами позвонков. В центре спинного мозга проходит канал (невроцель), продолжение желудочка головного мозга. Центральная часть спинного мозга состоит из серого вещества, периферическая – из белого. Спинной мозг имеет сегментное строение, от каждого сегмента, число которых соответствует количеству позвонков, с двух сторон отходят нервы.
Спинной мозг при помощи нервных волокон связан с различными отделами головного мозга, осуществляет передачу возбуждений нервных импульсов, также является центром безусловных двигательных рефлексов.
ОРГАНЫ ЧУВСТВ
Органы химической рецепции и осязания.Органы химической рецепции служат для получения информации о веществах, растворенных в воде, вкусе пищи. Они включают:
1) органы обоняния;
2) органы химической необонятельной рецепции. Органы обоняния (обонятельные мешки) расположены в носовой полости. У рыб обычно парные носовые отверстия (ноздри).
Ноздря разделена кожистым клапаном и имеет два отверстия, вода заходит в обонятельный мешок через переднее отверстие и выходит через заднее. Полость обонятельного мешка имеет складки (розетки) и выстлана слизистой оболочкой, связанной с нервными окончаниями. К органам обоняния подходят обонятельные нервы, отходящие от переднего мозга, и волокна тройничного нерва (рис.2).
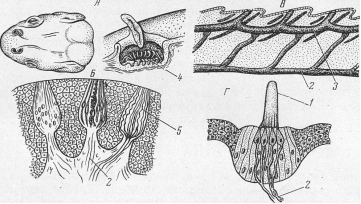
Рис. 2. Органы чувств рыб:
А — орган обоняния; Б — орган вкуса; В — боковая линия; Г — невромаст ; 1 — купула; 2 — нерв; 3 — канал боковой линии; 4 — обонятельная розетка; 5 — вкусовая почка.
Круглоротые имеют непарный орган обоняния (у миксин он сообщается с глоткой, у миног – нет). У миног ноздря ведет в длинный канал, задняя стенка которого образует обонятельную капсулу с чувствительными клетками. Канал продолжается до начала хорды, образуя питуитарный вырост. К верхней части этого выроста прилегает гипофиз. Обонятельный канал миног называют также назогипофизарным.
Движение воды (забор и выталкивание ее обратно) в обонятельной капсуле осуществляется за счет изменения объема питуитарного выроста. У миксин питуитарный вырост открывается отверстием в ротовую полость, поэтому у зарывшейся в ил минсины вода может поступать к жаберным мешкам через ноздри.
Рыбы имеют парные органы обоняния. У хрящевых рыб парные ноздри расположены обычно на брюшной стороне рыла. У всех рыб (за исключением двоякодышащих и кистеперых) ноздри с полостью глотки не сообщаются.
При помощи органов обоняния рыбы находят пищу, различают пол, физиологическое состояние рыб, ориентируются в пространстве. Некоторые виды очень чувствительны к запахам (акулы, проходные лососевые, налим, угорь и др.). Так, акулы способны распознавать запах крови на расстоянии до 2 км.
Рыбы чутко реагируют на сигналы опасности, на вещества, выделяемые из кожи при ранении. Реакция рыб различна: одни зарываются в ил, другие затаиваются, третьи выпрыгивают из воды и т.д.
Органы обоняния играют важную роль при миграциях рыб. Так лососевые рыбы запоминают запах (химический состав) реки, в которой они выклюнулись из икринки, и через несколько лет после нагула в море, возвращаются для размножения в родной водоем (явление хоминга – нахождение дома). Опыты с мечением личинок показали, что из 13 тыс. выловленных на нерестилищах рыб 34% вошли точно в те реки и ручьи, где они выклюнулись из икры, 65% в соседние и лишь 1% был пойман на значительном удалении от мест мечения.
Органы химической необонятельной рецепции воспринимают различные химические показатели среды (соленость, активная реакция среды, концентрация углекислоты и др.) и представлены:
1) вкусовыми почками (скопление чувствующих клеток);
2) вкусовыми клетками (клубковидные, кустиковидные, веретеновидные);
3) свободными нервными окончаниями.
У рыб центр химической необонятельной рецепции находится в
продолговатом мозгу.
Вкусовые почки и клетки расположены главным образом в слизистой полости рта, на усиках, жабрах, голове, лучах плавников.
Рыбы различают оттенки вкуса (сладкое, горькое, соленое, кислое). Острота вкуса связана с экологическими особенностями вида (пищевая специализация, тип местообитания, степень развития других рецепторов). Так, слепая мексиканская пещерная рыбка распознает раствор глюкозы при концентрации 0,005%.
Рыбы способны воспринимать тактильные (прикосновение, давление), болевые, температурные ощущения. Тактильные ощущения воспринимаются с помощью органов осязания. К ним относятся осязательные тельца (скопление чувствующих клеток), рассеянных по поверхности тела. Много осязательных точек расположено на голове, усиках и плавниках рыбы. Рыбы имеют невысокую болевую чувствительность из-за низкого уровня развития нервной системы.
Рыбы чувствительны к изменениям температуры. Температура воды воспринимается рыбами с помощью терморецепторов (свободных нервных окончаний), расположенных в поверхностных слоях кожи. Небольшие отклонения в температуре воды могут изменить пути миграций и сроки нереста рыб. Костные рыбы способны различать перепады температур в 0,4°С.
Электрические рецепторы и электрические органы рыб.По характеру вырабатываемых электрических импульсов рыб делят на:
1) неэлектрических (большинство других рыб).
2) слабоэлектрических (мормиры, гимнарх, гимнот и др.);
3) сильноэлектрических (электрические сом, угорь, скат);
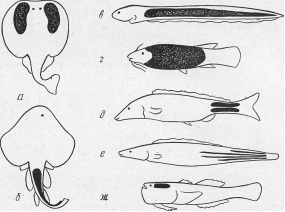
Рис.3. Расположение электрических органов у электрических рыб (по Гриффину и Новику):
а — электрический скат; б — морская лисица; в — электрический угорь;
г — электрический сом; д — слонорыл; е — гимнарх; ж — звездочет (выделенные участки на контурном изображении каждой рыбы представляют собой проекции электрических органов).
Электрические поля образуются вокруг тела любой рыбы. Неэлектрические рыбы создают слабые электрические импульсы 100–200 мкВ в результате нервно-мышечной деятельности. В стае электрические поля отдельных рыб суммируются и образуется общее биоэлектрическое поле, которое влияет на поведение и ориентацию рыб. Слабо- и сильноэлектрические рыбы имеют специализированными электрическими органами.
Рыбы воспринимают электрические поля по-разному. У слабо- и сильноэлектрических рыб (за исключением электрического сома и звездочета) имеются специальные электрорецепторы, которые входят в систему органов чувств боковой линии. У неэлектрических рыб (кроме хрящевых, некоторых осетровых, сомовых) электрорецепторы не обнаружены. Но они способны воспринимать электрический ток, действующий на другие рецепторы и свободные нервные окончания. Электрические органы рыб служат им для защиты, нападения на добычу и ориентации.
Электрические органы всех электрических рыб представляют собой парные, симметрично расположенные по бокам тела структуры, которые состоят из электрических пластинок, собранных в столбики (рис.3).
У ската электрические органы (до 25% массы рыбы) напоминают пчелиные соты. Один орган состоит приблизительно из 600 шестигранных призм, расположенных вертикально. В каждой призме насчитывается до 40 электрических пластинок, имеющих вид дисков, отделенных студенистой соединительной тканью. Каждая призма представляет собой своеобразную электрическую батарею.
У угря огромные электрические органы (около 30% массы тела) тянутся по бокам почти вдоль всего тела и также состоят из призм, но с горизонтальным расположением. В каждом органе насчитывают около 70 призм, каждая содержит около 6 тыс. электрических пластинок.
У сома парные электрические органы (около 25% массы тела) расположены под кожей вдоль тела и сходятся по средней линии спинной и брюшной сторон. В студенистом веществе электрических органов находится большое количество электрических пластинок (около 2 млн), расположенных поперек тела.
Нервными центрами электрических органов у рыб являются электрические доли продолговатого мозга и спинной мозг.
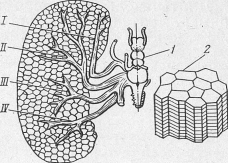
Рис. 4. Схема строения электрического органа ската:
I, II, III, IV — электрические нервы;
1 — электрические доли головного мозга; 2 — электрические столбики.
Основными элементами электрических органов у большинства рыб являются электрические пластинки (рис. 4). Электрическая пластинка имеет две стороны:
1) мембранную, или лицевую (к ней подходят нервные окончания);
2) обратную (к ней подходят кровеносные сосуды).
В момент возбуждения мембранная сторона пластинки становится электроотрицательной, а обратная – электроположительной. Электрические клетки в столбиках или призмах соединены последовательно (т.е. увеличивается общее напряжение), а ряды столбиков в электрических органах соединены параллельно (т.е. увеличивается общая сила тока).
Величина генерируемого напряжения различна. Обитатели пресных вод со слабой электропроводностью (электрические угорь и сом) генерируют ток высокого напряжения: угорь до 600 В при силе тока 1,2 А, сом до 350 В при силе тока в десятые доли ампера.
Обитатели морских вод с хорошей электропроводностью генерирует разряды меньшего напряжения, но высокой силы тока (электрический скат 40–60 В при силе тока 50–60 А).
Звездочеты и обыкновенные скаты занимают промежуточное положение между сильно- и слабоэлектрическими рыбами. Они имеют специализированные органы небольшого размера, которые расположены в хвостовой части тела (скат) или на голове (звездочет). Напряжение, создаваемое скатом-морской лисицей, составляет около 4 В.
Слабоэлектрические рыбы имеют небольшие хвостовые электрические органы, создаваемое напряжение – десятые доли вольта.
У круглоротых (морская минога и миксины) имеется слабый электрический орган на голове, способный создавать слабое электрическое поле напряжением до 1 милливольта. По изменению этого поля животные чувствуют приближение другого организма или препятствия.
Органы зрения.Глаза у большинства рыб расположены по бокам головы. Зрение у рыб монокулярное, т.е. каждый глаз видит самостоятельно (поле зрения по горизонтали 160–170°, по вертикали около 150°). У многих рыб хрусталик выступает из отверстия зрачка, что увеличивает поле зрения. Спереди монокулярное зрение каждого глаза перекрывается, и образуется бинокулярное (всего 15–30°). Основной недостаток монокулярного зрения неточная оценка расстояния (рис. 5)
У многих пресноводных рыб зрачок неподвижен, некоторые виды могут его сужать и расширять (угорь, камбалы, звездочет, хрящевые). Глаза большинства рыб не имеют век, у некоторых акул есть мигательная перепонка, у кефалей и некоторых сельдей развиваются жировые веки.
У рыб глаз включает три оболочки:
1) склера (наружная);
2) сосудистая (средняя);
3) сетчатка, или ретина (внутренняя).
Склера защищает глаз от механических повреждений, в передней части глаза образует прозрачную роговицу уплощенной формы. С помощью сосудистой оболочки осуществляется кровоснабжение глаза. В участке, где в глаз входит зрительный нерв, располагается характерная для рыб сосудистая железа. В передней части глаза сосудистая оболочка переходит в радужную, имеющую отверстие, – зрачок, в который выдается хрусталик.

Рис. 5.Строение глаза и поле зрения рыб
1 – склера; 1а, 1б, 1в – слои роговицы; 2 – сосудистая оболочка; 2а – пигментный слой сосудистой оболочки; 3 – радужина; 4 – хрусталик; 5 – задняя камера; 6 – передняя камера; 7 – кольцевая связка; 8 – сократительная мышца; 9 – серповидный отросток; 10 – сетчатка; 11 – зрительный нерв
Сетчатка включает:
1) пигментный слой (пигментные клетки);
2) светочувствительный слой (светочувствительные клетки: палочки и колбочки);
3) два слоя нервных клеток.
У большинства рыб в сетчатке имеются палочки и колбочки. Палочки функционируют в темноте и нечувствительны к цвету, колбочки воспринимают цвета.
Хрусталик в верхней части поддерживается связкой, а в нижней части он при помощи особой мышцы (колоколом Галлера) прикрепляется к серповидному отростку на дне глазного яблока, который имеется у большинства костистых рыб. Хрусталик у рыб шаровидный и своей формы не изменяет. Аккомодация (настройка на резкость) осуществляется не путем изменения кривизны хрусталика, а при помощи мышцы (колокол Галлера), которая подтягивает или удаляет хрусталик от сетчатки. Хрусталик имеет такую же плотность, как и вода, в результате чего свет, проходя через него, не преломляется и на сетчатке получается четкое изображение.
В зависимости от наличия светочувствительных клеток (палочек, колбочек) рыб подразделяют на:
1) сумеречных (в пигментном слое мало меланина, в сетчатке имеются только палочки);
2) дневных (в пигментном слое много меланина, в сетчатке палочки немногочисленны, колбочки крупные).
Рыбы воспринимают световые волны в 400–750 нм. Почти все рыбы (кроме сумеречных и большинства хрящевых) имеют цветное зрение и некоторые из них могут изменять окраску тела.
У рыб различная острота зрения. Обычно они видят предметы на расстоянии не более 10–15 м. Хрящевые рыбы являются наиболее дальнозоркими, так как способны сужать и расширять зрачок глаза.
Со снижением освещенности у одних видов размер глаз увеличивается, и они способны улавливать слабый свет (глубоководные рыбы – морской окунь, светящиеся анчоусы), у других – размер глаз уменьшается (налим, речной угорь). У ряда глубоководных и
пещерных рыб глаза отсутствуют.
В воздушной среде глазами рыбы почти не видят, у некоторых из них для этой цели в глазах имеются специальные приспособления.
У рыбы четырехглазки каждый глаз разделен горизонтальной перегородкой на две части. В верхней части глаза хрусталик упрощен, а роговица выпуклая, что позволяет видеть в воздушной среде.
Орган равновесия и слуха.Круглоротые и рыбы имеют парный орган равновесия и слуха, который представлен внутренним ухом (или перепончатым лабиринтом) и расположен в слуховых капсулах задней части черепа. Перепончатый лабиринт состоит из двух мешочков:
1) верхний овальный;
2) нижний круглый.
У хрящевых лабиринт разделен на овальный и круглый мешочки не полностью. У многих видов от круглого мешочка отходит вырост (лагена), представляющий собой зачаток улитки. От овального мешочка во взаимно перпендикулярных плоскостях отходят три полукружных канала (у миног – 2, у миксин – 1). На одном конце полукружных каналов имеется расширение (ампула). Полость лабиринта заполнена эндолимфой. От лабиринта отходит эндолимфатический проток, который у костистых рыб заканчивается слепо, а у хрящевых сообщается с наружной средой (рис. 6).
Внутреннее ухо имеет волосковые клетки, которые являются окончаниями слухового нерва и расположены участками в ампулах полукружных каналов, мешочках и лагене. В перепончатом лабиринте есть слуховые камешки, или отолиты. Они располагаются по три с каждой стороны: один, самый крупный, отолит – в круглом мешочке, второй – в овальном, третий – в лагене. На отолитах хорошо видны годовые кольца, по которым у некоторых видов рыб определяют возраст (корюшка, ерш и др.).
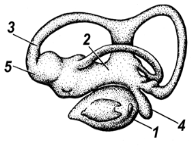
Рис. 6. Орган равновесия и слуха
1 – круглый мешочек; 2 – овальный мешочек; 3 – полукружные каналы; 4 – зачаток улитки; 5 – ампулы полукружных каналов
Верхняя часть перепончатого лабиринта (овальный мешочек с полукружными каналами) выполняет функцию органа равновесия, нижняя часть лабиринта воспринимает звуки. Любое изменение положения головы вызывает движение эндолимфы и отолитов и раздражает волосковые клетки.
Рыбы воспринимают в воде звуки в диапазоне от 5 Гц до 15 кГц, звуки более высоких частот (ультразвуки) рыбами не воспринимаются. Рыбы воспринимают звуки также и с помощью органов чувств системы боковой линии. Чувствительные клетки внутреннего уха и боковой линии имеют сходное строение, иннервируются ветвями слухового нерва и относятся к единой акустиколатеральной системе (центр в продолговатом мозгу). Боковая линия расширяет диапазон волн и позволяет воспринимать низкочастотные звуковые колебания (5–20 Гц), вызываемых землетрясениями, волнами и т.д.
Чувствительность внутреннего уха повышается у рыб с плавательным пузырем, который является резонатором и рефлектором звуковых колебаний. Соединение плавательного пузыря с внутренним ухом осуществляется при помощи Веберова аппарата (система 4 косточек) (у карповых), слепых выростов плавательного пузыря (у сельдевых, тресковых) или особых воздушных полостей. Наиболее чувствительными к звукам являются рыбы, имеющие Веберов аппарат.
При помощи плавательного пузыря, связанного с внутренним ухом, рыбы способны воспринимать звуки низких и высоких частот.
Органы чувств системы боковой линии.Строение и расположение органов чувств системы боковой линии специфичны для каждого вида рыб. Система боковой линии (или сейсмосенсорная система) включает обычные и ампулярные органы. К обычным органам относятся невромасты (или чувствующие почки), которые представляют комплекс чувствительных клеток с волосками на конце. У многих рыб невромаст образует студенистый выступ (купулу), куда входят волоски чувствительных клеток. Купулы легко колышутся под действием токов воды. Невромасты располагаются на поверхности тела в углублениях. Они позволяют рыбам ориентироваться в воде и воспринимать звуки низких частот. Клетки невромастов иннервируются боковой ветвью блуждающего нерва.
У круглоротых органы чувств системы боковой линии пред-
ставлены многочисленными мелкими бугорками на коже, особенно на голове. В каждом бугорке имеется желобок, на дне которого находится невромаст.
У хрящевых рыб имеются невромасты, расположенные в открытых (у примитивных акул и химер) и закрытых каналах (у других рыб); пузырьки Сави (Савишевы бляшки) – невромасты, прикрытые видоизмененной плакоидной чешуей (у электрических скатов); спиракулярные органы, представленные невромастами, поверхности брызгальца.
У осетровых имеются невромасты, расположенные в каналах и коротких костных трубочках кожи, и спиракулярный орган.
У большинства костистых рыб невромасты находятся в замкнутых каналах, которые тянутся вдоль туловища, образуя боковую линию, заходят на голову. Каналы боковой линии и головы заполнены слизью, в их полость выступают купулы чувствующих почек. С наружной средой каналы соединяются через отверстия в чешуе (прободенные чешуйки).
Ампулярные органы (или ампулы Лоренцини) относятся к электрорецепторам. Они расположены в головном отделе рыб, имеются только у пластиножаберных (акулы, скаты). Ампулярные органы имеют вид капсул, которые погружены в кожу. От капсул отходят трубочки, открывающиеся отверстиями на поверхности кожи. Капсулы заполнены желеобразным веществом, на их стенках расположены чувственные клетки с волосками на вершине. К каждой капсуле подходят нервные окончания.
К органам чувств системы боковой линии относятся также электрорецепторы, которые имеются почти у всех электрических рыб и у некоторых неэлектрических костистых. Они сходны по строению с чувствительными клетками системы боковой линии, но не имеют волоска.
Дата добавления: 2016-05-25; просмотров: 79811;