Симпатическая нервная система
Преганглионарные нейроны симпатической нервной системы расположены в боковых рогах, (сегменты С8—L2 —центр Якобсона). Аксоны преганглионарных нейронов синаптически связаны с нейронами симпатических ганглиев, расположенных экстраорганно (большинство в брюшной полости). От нейронов симпатических ганглиев идут постганглионарные волокна, иннервирующие все органы и ткани организма (см. рис. 81). Сосуды и потовые железы иннервируются от всех симпатических преганглионарных нейронов спинного мозга. В последнем шейном и двух первых грудных сегментах спинного мозга расположен спиноцилиарный центр, иннервирующий гладкие мышцы глаза, в том числе мышцу, расширяющую зрачок. Эффекторные нейроны этого пути лежат в краниальном шейном симпатическом ганглии. От 2—4-го грудного сегмента спинного мозга отходят симпатические волокна, иннервирующие слюнные железы. В Тh1—Тh5 расположены преганглионарные симпатические нейроны, иннервирующие сердце. Ганглионарные клетки этого пути лежат в основном в звездчатом ганглии или, реже, в узлах пограничного симпатического ствола.
Медиаторы и рецепторы:
Медиаторам и постганглионарных симпатических волокон являются норадреналин (около 90%), адреналин (около 7%), дофамин (около 3%). Синтезируется норадреналин из аминокислоты тирозина, в основном в терминальных симпатических окончаниях (только около 1% — в теле нейрона). Поскольку в окончаниях симпатических ганглионарных нейронов выделяется норадреналин, эти нейроны называют адренергическими. Катехоламин, как и другие медиаторы, выделяется при возбуждении нервного окончания путем везикулярного экзоцитоза, реализуемого с помощью Са2+, который проникает в пресинаптические нервные окончания через потенциалзависимые Са-каналы. Выведение Са2+ из нервного окончания после его возбуждения осуществляется Са-насосом.
При возбуждении постганглионарных симпатических волокон вместе с норадреналином выделяются также комедиаторы (нейропептиды — нейропептид Y, соматостатин, метэнкефалин), АТФ, играющие модулирующую роль.
Инактивирование медиатора, выделившегося в синаптическую щель, осуществляется за счет обратного его захватывания (75-80%) пресинаптической мембраной. Вернувшиеся в пресинаптическое окончание молекулы медиатора могут снова упаковываться в пузырьки для последующего использования. Не попавший в пузырьки норадреналин разрушается ферментами, локализованными в пресинаптической терминали, — МАО и катехол-О-метилтрансферазой. Часть норадреналина, выделившегося в синаптическую щель и не связавшегося с пост- и пресинаптическими рецепторами, диффундирует в интерстиций и кровь.
Эффекторные рецепторы. Адренорецепторы являются гликопротеидами, они имеют два типа: α- и β. Это деление рецепторов основано на их чувствительности к различным фармакологическим препаратам: α-адренорецепторы блокируются фентоламином, а β-адренорецепторы — пропранололом. Оба типа рецепторов подразделяются на два подтипа: α1- и α2-адренорецепторы, β1- и β2- адренорецепторы. В большинстве органов, реагирующих на катехоламины, содержатся α- и β-адренорецепторы, причем одна гладкомышечная клетка может иметь оба рецептора (см. табл. 10).
Иннервация адренорецепторов. Симпатическую иннервацию имеют преимущественно α1- и β2-адренорецепторы. Они активируются в основном медиатором норадреналином. Адренорецепторы α2 и β2 преимущественно не имеют иннервации, они расположены вне синапсов на клетках-эффекторах и активируются циркулирующим в крови адреналином и норадреналином, диффундирующим из синаптической щели после его выделения пресинаптическими окончаниями. Адренорецепторы α2 и β2 обнаружены также на пресинаптических окончаниях (пресинаптические рецепторы), где они регулируют выделение медиатора. Прямая связь посредством синапса имеет место в сердце, в жировой ткани, в сосудах и в ряде гладкомышечных органов (мышцы зрачка, матки, пиломоторы, семявыносящий проток, кишечник). Непрямой тип регуляции, как известно (В.А. Говырин), осуществляется в большинстве органов — скелетных мышцах, эндокринных железах, большинстве экзокринных желез. В этих органах медиатор из сосудистых симпатических сплетений диффундирует к местным, окружающим сосуд, клеткам либо разносится кровью.
Эффекты активации α- и β- адренорецепторов зависят от преобладания каждого из них в данном органе и от различной чувствительности α- и β-рецепторов к медиатору.
При возбуждении симпатических (адренергических) нервных волокон активация α1 - и α2- адренорецепторов клеток-эффекторов в разных органах вызывает различные реакции. В частности, сосуды суживаются, моторика ЖКТ угнетается (см. табл. 10). Если же симпатический ствол раздражается в грудной полости, то в регистрируются не тормозные, а стимуляторные реакции желудка и двенадцатиперстной кишки, что осуществляются, с помощью серотонинергических нервных волокон, имеющихся в составе симпатических стволов.
Таким образом, можно выделить серотонинергический отдел в составе ВНС (если в основу классификации отделов ВНС положить медиаторный принцип).
Схема вегетативной эфферентной иннервации (на примере желудка и тонкой кишки) представлена на рис. 87.
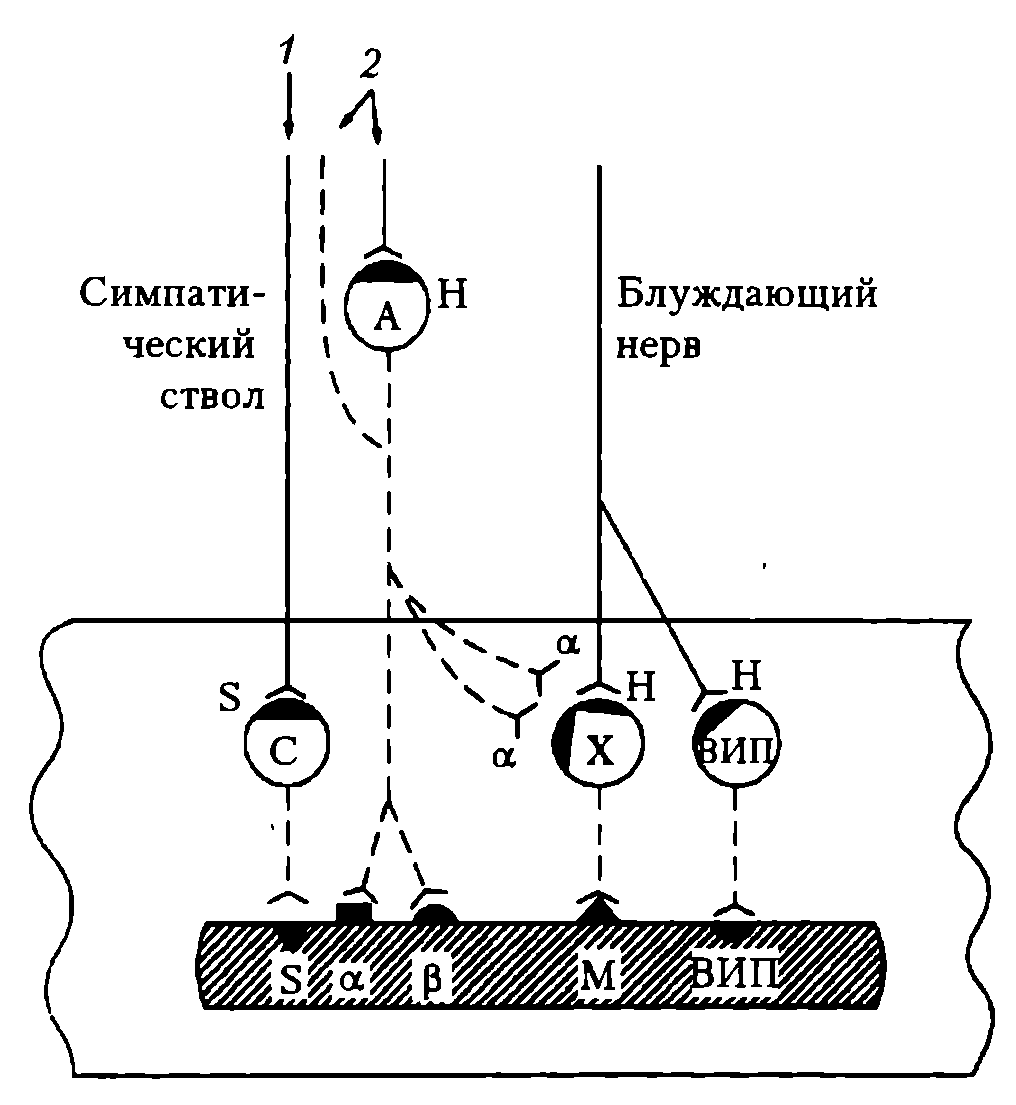
Рис. 87. Функциональная организация эфферентного отдела ВНС (на примере желудка и тонкой кишки): Нейроны: А – адрен-, С – серотонин-, Х – холин-, ВИПергические, и соответствующие их медиаторам α-, β-, S-, H-, M- и ВИП-рецепторы (ВИП – вазоактивный интестинальный пептид). Нервные волокна: 1 – серотонинергические, 2 – симпатические, сплошная линия – преганглионарные; пунктирная линия - постганглионарные
Активация β-адренорецепторов также вызывает разнонаправленные эффекты в различных органах: в ЖКТ она угнетает сократительную активность, в сердце (β1-рецепторы) — увеличивает частоту и силу сердечных сокращений.
Механизм действия катехоламинов, как и других медиаторов, зависит от класса эффекторных рецепторов.
Рецепторы, с помощью которых реализуется действие медиаторов нервных окончаний на клетки, делятся на два класса: ионотропные и метаботропные (рис. 88).
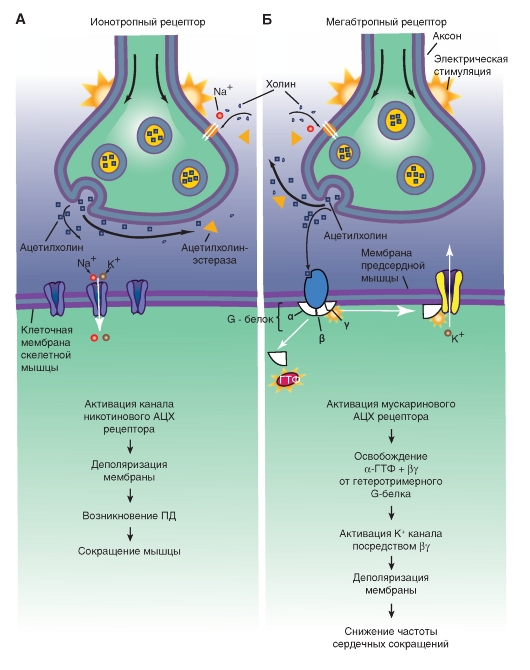
Рис. 88. Схема показывает механизм работы ионотропного (А) и метаботропного (Б) рецепторов постсинаптической мембраны при воздействии медиатора (Камкин, 2010)
Действие нейромедиатора ионотропных рецепторов ведет непосредственно к активации их ионных каналов. Действие нейромедиатора метаботропных рецепторов ведет к активации им вторых посредников, активирующих ионные каналы.
Эффекты гормонов реализуются в основном посредством метаботропных рецепторов, которые локализуются на клеточной мембране или в протоплазме.
Различия в механизме действия медиаторов связаны с тем, что у большинства постсинаптических рецепторов белок не является ионным каналом. Действие на них медиатора вызывает каскад химических реакций, в результате которых соседние ионные каналы активируются с помощью вторых посредников. Большинство рецепторов связано с G-белками. G-белки — ГТФ-связывающие белки. Активированный С-белок взаимодействует с помощью вторых посредников с ферментами. С G-белком связаны α- и β-адрено-, М-холино-, серотонино-, аденозино-, обонятельные, зрительные (родопсин) рецепторы, а также рецепторы большинства пептидных гормонов.
При передаче сигнала от нервных волокон посредством и метаботропных и ионотропных рецепторов к другим клеткам в последних вслед за электрофизиологическими процессами (деполяризация, гиперполяризация) активируются биохимические процессы. В первую очередь, это ресинтез АТФ, израсходованной на работу ионных помп, постоянно поддерживающих градиенты ионов внутри и вне клетки и усиливающих свою работу для устранения ионных сдвигов, произошедших при деполяризации или гиперполяризации клеток (мышечных, нервных, секреторных). Кроме того, в мышечных и секреторных клетках энергия АТФ расходуется на их сокращение, в нервных клетках запускаются биохимические процессы и структурные изменения (синтез белка), обеспечивающие хранение информации (память). Гормоны же свое влияние реализуют с помощью мембранных или внутриклеточных рецепторов, изменяя интенсивность метаболических процессов с помощью вторых посредников без электрофизиологических явлений.
Фосфорилирование белка — один из главных способов его активации с помощью протеинкиназ. Все клетки содержат протеинфосфотазы и дефосфорилирующие белки, обеспечивающие обратное действие — убирающие эффект фосфорилирования белков.
Протеинкиназы, зависимые от вторых посредников: цАМФ-зависимая протеинкиназа, протеинкиназа-С, кальмодулинзависимые протеинкиназы (один из путей реализации эффекта — связывание белка кальмодулина с кальцием, комплекс регулирует активность многих белков).
Все α- и β-адренорецепторы являются метаботропными. Эффекты α1- адренорецепторов реализуются с помощью вторых посредников — инозитолтрифосфата и диацилглицерола (ИФ3/ДАГ). При активации α2-адренорецепторов снижается скорость синтеза цАМФ в результате активации G-белка.
При действии норадреналинана β-адренорецепторы эффект реализуется с помощью аденилатциклазы и накопления цАМФ внутри клетки.
При избытке норадреналина формируется снижение чувствительности клеток к медиатору (десенситизация), а при недостатке норадреналина наблюдается повышение их чувствительности (сенситизация).
Преганглионарные симпатические волокна образуют возбуждающие холинергические синапсы также и на клетках мозгового слоя надпочечников. Выделение катехоламинов из мозгового вещества надпочечников регулируется исключительно нервными влияниями (перерезка преганглионарных симпатических волокон прекращает секрецию катехоламинов). При возбуждении преганглионарных симпатических волокон у человека из надпочечников в кровоток обычно выбрасывается смесь катехоламинов, состоящая из адреналина (80—90%) и норадреналина (10-20%).
Симпатоадреналовая система активирует деятельность организма, является пусковым звеном стрессорных эмоционально окрашенных реакций, мобилизует защитные силы организма, обеспечивает выход крови из кровяных депо, поступление в кровь глюкозы, ферментов, усиливает метаболизм тканей, увеличивает расход энергии.
Дата добавления: 2016-02-16; просмотров: 1951;