Клинические проявления поражений вирусом простого герпеса
Оба вируса простого герпеса (ВПГ 1-го типа и ВПГ 2-го типа) вызывают аналогичные поражения, локализация которых специфична и связана с типом возбудителя.
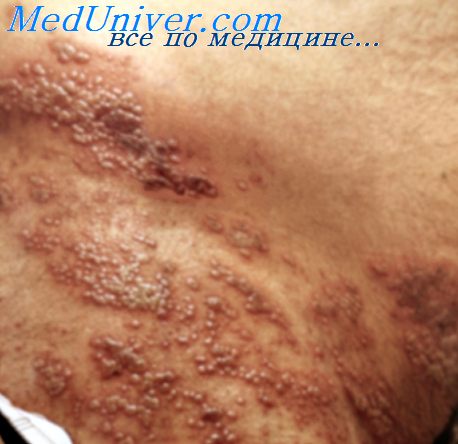
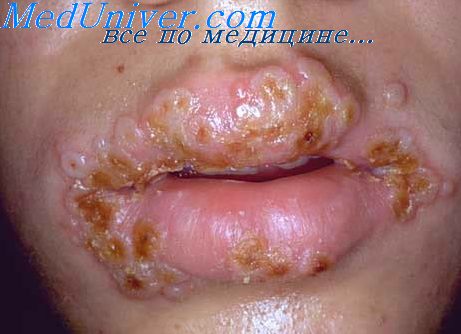
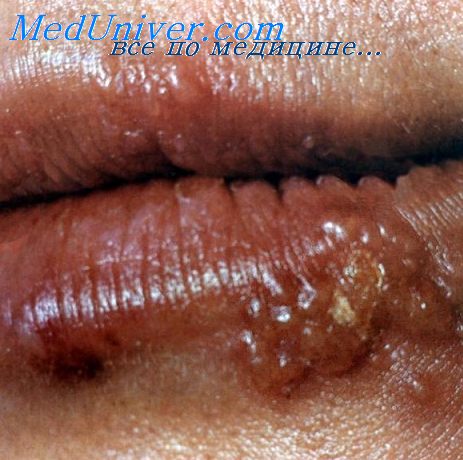
Герпетический гингивостоматит. Основной возбудитель— ВПГ 1-го типа. Для первичной инфекции характерны поражения многослойного эпителия красной каймы губ, часто сопровождающиеся дисфагией и лихорадкой. Выздоровление обычно наступает через 7-10 сут. Рецидивирующие поражения обычно наблюдают после переохлаждения. Рецидивы часто сопровождаются недомоганием, лихорадкой; иногда они носят генерализованный характер.
Генитальный герпес. Основной возбудитель — ВПГ 2-го типа. Типичны характерные поражения наружных половых органов, создающие дискомфортные ощущения; более тяжёлые случаи сопровождаются общим недомоганием и лихорадкой. Поражения обычно исчезают через 10-14 дней, но часто рецидивируют.
Герпетический менингознцефалит чаще вызывает ВПГ 2-го типа, обычно протекает стёрто. Патогенез включает прогрессирующую демиелинизацию нервных волокон; первоначально у пациентов отмечают психические, а затем и неврологические нарушения.
Герпетический кератит. Основной возбудитель— ВПГ 1-го типа. Поражения могут быть первичными и рецидивирующими; последние способны приводить к необратимой потере зрения вследствие развития язв роговицы, особенно в области зрачка.
Герпес новорожденных — тяжёлое генерализованное поражение; инфицирование происходит при прохождении через родовые пути. Заболевание чаще вызывает ВПГ 2-го типа.
Семейство иридовирусы является обширным и сложным. Оно включает три рода вирусов: ранавирусы (вирус лягушек 3), лимфоцистивирусы и род без названия (вирус золотистых рыб 1).
Несколько представителей семейства иридовирусы связаны с болезнями рыб и амфибий. Наиболее известный вирус лимфоцистита рыб вызывает опухолеоб-разные изменения на коже. Он вызывает заболевание более чем у 90 различных видов морских и пресноводных рыб и является важным патогеном.
Иридовирусы — оболочечные вирусы диаметром 300 нм. Иридовирусы позвоночных морфологически подобны вирусу АЧС. Иридовирусы имеют сложный икосаэдрический капсид диаметром 130—170 нм. Геном представлен одной линейной молекулой двухцепочной ДНК размером 95000-190000 пар оснований. Иридовирус москитов имеет геном 440000 пар оснований и является наиболее крупным геномом ДНК вирусов. Для транскрипции и репликации ДНК необходимо ядро клетки, но некоторые ДНК синтезируются и включаются в вирионы в цитоплазме. Концы генома иридовирусов отличаются от концов генома вируса АЧС циркулярными перестановками, концевой избыточностью и содержанием метилированных оснований, подобно ДНК бактерий.
Иридовирусы позвоночных размножаются в различных культурах клеток амфибий и млекопитающих при температуре 12-32°С. Их репликация подобна вирусу АЧС. Однако их геном не кодирует РНК полимеразу, а использует клеточную РНК полимеразу II, которая модифицируется структурными белками для преимущественного синтеза вирусной мРНК. Другое отличие заключается в том, что первая стадия репликации ДНК иридовирусов происходит в ядре, а вторая - в цитоплазме с образованием конкатемеров в 10 и более превышающих размер вирусного генома. Иридовирусы позвоночных, также как асфаровирусы, образуют крупные параклисталлические структуры вирионов в цитоплазме инфицированных клето.
Для вирусов этого семейства характерно исключительное разнообразие и широкий круг естественных хозяев (от высших приматов до грибов).
Семейство герпесвирусов (ГВ) насчитывает более 100 представителей, объединенных в одно семейство исходя из общности их структурных характеристик. Их обнаруживают почти у всех исследуемых животных. Особенно многочисленные представители герпесвирусов циркулируют в человеческой популяции. Герпес (от греческого herpes - ползучий) - одна из самых распространенных вирусных инфекций человека. Герпесвирусам принадлежит важная роль в патологии сельскохозяйственных животных и птиц, а также промысловых рыб.
По крайней мере одно из главных заболеваний трех видов домашних животных вызывается герпесвирусом, в их числе инфекционный ринотрахеит крупного рогатого скота, болезнь Ауески свиней и болезнь Марека кур. Многие герпесвирусы охарактеризованы частично, а у 19 из них, включая наиболее патогенные герпесвирусы, геном был полностью секвенирован. Очень немногие семейства вирусов животных имеют такое разнообразие представителей, как герпесвирусы.
Персистентная инфекция герпесвирусами сопровождается периодическим или постоянным выделением вируса. При альфагерпесвирусных болезнях множественные копии вирусной ДНК существуют как эписомы или, реже, интегрированы в хромосомальную ДНК латентноинфицированных нейронов. Латентные геномы исключительно «молчаливы», за исключением образования латентносвязанных транскриптов. Неизвестно, чтобы эти РНК-транскрипты кодировали какой-либо протеин (герпесвирус 1 КРС является исключением).
Механизм установления и поддержания латентной инфекции еще не известен. Реактивация обычно связана со стрессом, половыми инфекциями, транспортировкой, охлаждением, перенаселением. Реактивация ГВ домашних животных обычно не наблюдается.
Герпесвирусы способны оставаться в латентном состоянии в организме естественных хозяев. В клетках, содержащих латентный вирус, вирусный геном находится в форме циркулярных молекул, и только небольшая часть вирусных генов экспрессируется.
Латентные геномы остаются способными к репликации и вызывают заболевание после реактивации. Молекулярные механизмы, ведущие к реактивации латентного состояния, полностью неизвестны и могут отличаться от механизма реактивации других вирусов. Латентность отличается от хронической инфекции отсутствием потомства инфекционного вируса. Наоборот, способность к реактивации отличает латенцию от абортивной инфекции.
Основным фактором, определяющим эпидемиологические или эпизоотологические особенности герпесвирусов, является способность оставаться в латентной форме в чувствительных и вегетативных ганглиях. Вирусный геном может длительное время персистировать в клетках без репликации вируса. Опасность представляет реактивированный вирус, локализованный в клинически выраженных очагах поражения при возвратной инфекции или выделяемый из клинически инапарантных очагов поражения. После первичного инфицирования многие герпесвирусы могут длительно персистировать в организме естественного хозяина, периодически давая рецидивы болезни.
У теплокровных животных реактивация вирусов происходит под воздействием различных факторов (стрессы, гормональные сдвиги, травмы, интеркуррентные заболевания, ультрафиолетовое облучение, лихорадка) и, вероятно, связана с подавлением Т-клеточного иммунитета. Методом гибридизации in situ транскрипты специфической вирусной РНК выявлены в ядрах нейронов латентноинфицированных животных. Однако у разных представителей герпесвирусов механизм латенции и реактивации может быть различным. Некоторые герпесвирусы вызывают неоплазмы у их естественных хозяев, экспериментальных животных или у тех и других, а также иммортализацию клеточных культур. Несмотря на большое разнообразие естественных хозяев, герпесвирусы объединяет поразительное сходство. Принадлежность вирусов к этому семейству определяется на основе морфологии вирионов и свойств ДНК. Несмотря на то, что различные герпесвирусы не удается дифференцировать по морфологии, не существует других семейств вирусов, представители которых характеризовались бы столь большой вариабельностью. Герпесвирусы, однако, легко дифференцировать на основе биологических свойств, иммунологической специфичности, по размеру вирионов и первичной структуре генома. Для этих вирусов характерно почкование созревших вирионов через внутреннюю мембрану клеточного ядра.
Герпесвирусы могут передаваться от одного поколения хозяев к другому благодаря персистенции, часто латентной инфекции, при которой вирус периодически реактивируется и выделяется из организма. При некоторых герпесвирусных инфекциях выделение вируса может быть фактически непрерывным. Молекулярное филогенетическое изучение подтверждает, что, за небольшими исключениями, каждый вирус является уникальным, потому что он происходит от своего видового хозяина. Латентность позволяет герпесвирусам быть пожизненными даже в очень небольшой изолированной группе хозяев.
Попытки разработать критерии классификации герпесвирусов внутри семейства пока не увенчались успехом. Существующая классификация основана на их естественном паразитизме.
В зависимости от биологических свойств герпесвирусы делятся на 4 подсемейства, 3 из которых имеют названия альфа-, бета-, гаммагерпесвирусы и одно - без названия, объединяющие герпесподобные вирусы.
Подсемейство альфагерпесвирусы включает герпесвирусы 1 и 3 человека, герпесвирус 1 кур и вирус болезни Марека. Большинство альфагерпесвирусов размножаются быстро (цикл репликации меньше 24 ч) и вызывают латентную инфекцию первоначально в сенсорных ганглиях. Подсемейство бетагерпесвирусы представлено цитомегаловирусами многих видов млекопитающих. Прототипным вирусом является герпесвирус человека 5 — цитомегаловирус человека. Вирусы этого подсемейства размножаются медленно (цикл репликации более 28 ч) и вызывают лизис клеток спустя несколько дней. Они могут оставаться латентными в секреторных железах, лимфоретикулярных тканях, почках и других органах. Подсемейство гаммагерпесвирусы представляет лимфотропные вирусы. Прототипом является герпесвирус человека 4 — вирус Эпштейн-Барр. Гаммагерпесвирусы обладают узким хозяинным спектром и находятся латентно в лимфоцитах (лимфома Беркитта и др.), некоторые представители вызывают инфекцию с разрушением клеток эпителия и фибробластоидных клеток.
Вирионы герпесвирусов имеют диаметр около 120—150 нм и состоят из 4 структурных компонентов: сердцевины (ядра), содержащей линейную двуцепочечную ДНК размером 125—235 тпн, икосаэдрического нуклеокапсида диаметром 100-110 нм, содержащего 162 капсомера; аморфного материала (тегумент), окружающего капсид и оболочки с небольшими гликопротеиновыми выступами (пепломерами) на поверхности.
ДНК-геном закручен вокруг волокнистого ядра, имеющего вид катушки, которая прикреплена волокнами к внутренней поверхности окружающего капсида. Вирусный геном в соответствующих экспериментальных условиях обладает инфекционностью.
Сердцевина вириона окружена икосаэдрическим капсидом, содержащим 162 частично полых капсомера (12 пентамеров и 150 гексамеров). Капсомеры высотой 12,5 нм и диаметром 9,5 нм имеют внутренний осевой канал диаметром 4 нм, простирающийся внутрь на половину длинной оси капсомера.
Слой, лежащий между оболочкой вириона и нуклеокапсидом (тегумент), содержит глобулярный материал, часто распределяемый ассиметрично, и выражен неодинаково у разных герпесвирусов. У вируса герпеса простого он развит слабо, тогда как у вирусов болезни Марека (БМ) и чумы уток (ЧУ) он развит отчетливо и его наличие обусловливает крупный размер вирионов. На поверхности герпесвирусов находится двухслойная оболочка с многочисленными небольшими гликопротеиновыми пепломерами, образованными главным образом за счет «бляшек» измененной ядерной мембраны инфицированной клетки.
Количество гликопротеинов в оболочке герпесвирусов варьирует. В оболочке вируса простого герпеса их не менее 11. Число копий индивидуальных гликопротеинов может превышать 1000 на вирион. Содержание полипептидов в вири-оне точно не известно и различается у разных герпесвирусов. В общем, оно колеблется в пределах 35—45 полипептидов.
Все герпесвирусы кодируют широкий набор ферментов, связанных с метаболизмом нуклеиновых кислот, синтезом ДНК и процессингом белков. Синтез вирусных ДНК и сборка вирионов происходят в ядре клетки. Капсиды покрываются оболочкой, проходя через мембрану ядра. Образование инфекционного вирусного потомства сопровождается деструкцией зараженных клеток. Вероятно, что некоторые кодируемые вирусом белки играют заметную роль в патогенезе и иммунитете при герпесвирусной инфекции, не являясь необходимыми для репликации вируса в культуре клеток. Считают, что геномы герпесвирусов в латентном состоянии в клетках хозяина существуют в циркулярной эписомальной (экстрахромосомальной) форме.
Репликация герпесвирусов наиболее детально изучена на примере герпесвируса 1 человека (вирус простого герпеса 1). После прикрепления вириона путем связывания пепломерных гликопротеинов оболочки с рецепторами клетки, одним из которых является гепаринсульфатпротеогликан, нуклеокапсид входит в цитоплазму путем слияния оболочки вириона с мембраной клетки путем эндофагоцитоза. Затем ДНК-белковый комплекс освобождается от нуклеокапсида и быстро останавливает синтез макромолекул клетки.
Геном ВПГ-1 (153 тпн) содержит три области инициации-репликации. Он практически кодирует все гены, необходимые для репликации, кроме ДНК-лигазы и топоизомеразы. На первом этапе репликации функционирует ДНК-полимераза а, а затем функционирует вирусная ДНК-полимераза.
Три класса мРНК — а, (3 и 7-транскрибируются последовательно клеточной РНК полимеразой II. Таким образом а (ранние) РНК в последующем функционируют как мРНК и транслируются в а-белки, которые вызывают транскрипцию (3 (ранних) мРНК, трансляция которых приводит к синтезу (3 (ранних) белков и подавляет дальнейшую транскрипцию а мРНК. Затем начинается репликация вирусной ДН К с использованием тех же самых аир протеинов, так же как протеинов клетки. Транскрипционная программа затем возобновляется, и в результате появляются 7 (поздние) мРНК, которые транслируются в 7 протеины. Более 70 белков, кодируемых вирусом, образуются в цикле его репродукции, многие из а и (3 белков являются ферментами и белками, связанными с вирусной ДНК, тогда как 7 белки являются структурными вирионными компонентами. Сложный контроль регуляции экспрессии осуществляется на уровне транскрипции и трансляции. Вирусная ДНК реплицируется в ядре, и вновь синтезированная ДНК упаковывается в преформированные незрелые капсиды. Созревание вирионов связано с вхождением ДНК в нуклеокапсиды и связыванием нуклеокапсидов с измененным внутренним слоем оболочки ядра и последующим обволакиванием вирионов при почковании. Зрелые вирионы накапливаются внутри вакуолей в цитоплазме и освобождаются экзоцитозом или при цитолизе. Вирусспецифические белки обнаружены также в цитоплазматической мембране, где они участвуют в клеточном слиянии и могут в качестве Fc-рецепторов выступать мишенями в иммунном цитолизе. Герпесвирусная инфекция сопровождается образованием внутриядерных телец включений, которые обычно обнаруживают в фиксированных окрашенных препаратах.
Геном вируса кодирует синтез по меньшей мере четырех гликопротеинов (gB, gC, gD, gE). Имеет место посттрансляционное расщепление или другие модификации вирусных белков. Белок gB обладает фузогенной активностью и ответственен за проникновение вируса в клетку; gD — основной протективный антиген.
Белки gC и gE не играют существенной роли в репродукции вируса, но помогают ему ускользнуть от иммунной системы организма. В отсутствие репликации вирус редко вызывает сплавление клеток, а при наличии специфических условий — агглютинацию эритроцитов. Оболочка вириона играет существенную роль в антигенности и инфекционности вируса. Гликопротеины вирусной оболочки ответственны за прикрепление вирусной частицы к клетке, проникновение вируса в клетку, за индукцию нейтрализующих антител и образование иммунитета. Нейтрализующие антитела реагируют с главными вирусными гликопротеинами, находящимися в оболочке вириона. Мембранные антигены, индуцированные в зараженных клетках, содержат по крайней мере три гликопротеина, обнаруженных в структуре вириона. Эти данные послужили основанием для приготовления вакцин из вирусспецифических компонентов. Однократное введение вакцины из субъединиц приводило к активизации гуморальных и клеточных факторов иммунитета у мышей и кроликов, повышало их резистентность к экспериментальному заражению и снижало количество клеток ганглия тройничного нерва, несущих геном вируса.
Основным протективным антигеном вируса простого герпеса типа I и II (ВПГ-1, ВПГ-2) является гликопротеин Д (gD), и в меньшей степени - гликопротеин B(gB).
Очищенный gD ВПГ-1 или ВПГ-2 защищает мышей от летальной дозы как ВПГ-1, так и ВПГ-2; стимулирует образование ВН-антител, пассивное введение которых защищает мышей от летальной инфекции ВПГ; стимулирует комплементзависимую и опосредованную антителами цитотоксичность. Комбинация обоих гликопротеинов (gD и gB) усиливает протективную активность препарата. В четырех антигенных участках gD ВПГ-1 различают по крайней мере шесть эпитопов. Антитела к большинству из них нейтрализуют вирус без участия комплемента и подавляют проникновение вируса в клетку. Нейтрализация вируса и индукция протективного иммунитета приводили к активизации гуморальных и клеточных факторов иммунитета у мышей и кроликов, повышали их резистентность к экспериментальному заражению и снижали количество клеток ганглия тройничного нерва, несущих геном вируса. Нейтрализация вируса и индукция протективного иммунитета связаны с аминокислотными остатками 233-259, а адсорбция вируса - с областью в пределах первых 182 аминокислот. Точечные мутации в сайте нейтрализации gD могут сделать его нечувствительным к действию ВН-антител.
Слияние вирусной и клеточной оболочек ВПГ-1 определяет gB. За аналогичные функции вируса Эпштейн-Барра ответственны два гликопротеина (220/200 и 350/300 кД), расположенные на поверхности вириона. Эти белки вызывали у кроликов образование антител, реагирующих с одними и теми же белками на поверхности вирионов и инфицированных вирусом клеток. Вакцина, содержащая гликопротеин 340 кД, вызывала иммунитет у обезьян. В нейтрализации цитомегаловируса и выработке нейтрализующих антител основная роль принадлежит гликопротеинам 86 и 130/55 кД.
В оболочке вируса псевдобешенства (ВПБ) различают три основных гликопротеина (gE, gB, gX), кодируемых u-сегментом вирусного генома. При иммунном ответе свиней на инфекцию защитные антитела формируются к двум гли-копротеинам: gE и gB. Гликопротеин gE состоит из двух субъединиц с массой около 80 кД. Он не является существенным для репликации, поскольку известны вакцинные штаммы с делениями в этой области генома. N-концевой фрагмент gE содержит высокоантигенный домен, способный вызвать образование НВ-антител и защитный эффект. Делеция по гену gE может привести к снижению иммунногенности, чего не наблюдается при удалении гена gX. Предшественники gII и gIII имеют соответственно молекулярную массу 110 и 60 кД. За слияние вирусной и клеточной оболочек и проникновение вируса в клетку ответственен gB. Антигенного дрейфа в gB и gE не обнаружено.
Однако основную роль в индукции синтеза нейтрализующих антител ВПБ, проявляющих активность в отсутствие комплемента, играет gB. Являясь мультифункциональным белком, он, кроме того, играет важную роль в адсорбции и эффективной репликации вируса в культуре клеток. Антитела к gIII обладали более выраженным защитным эффектом у мышей (90-100%) и свиней (84-100%), чем к gII. Гликопротеин gIII играет определяющую роль в клеточном иммунитете, поскольку является главным антигеном-мишенью для вирусспецифических цитотоксических Т-лимфоцитов. Кроме gill, заметная роль в защите вакцинированных животных принадлежит g50 кД, который также является основной мишенью ВН-антител. Следует отметить, что пассивная защита поросят не предотвращала репликацию и экскрецию ВПБ в назофаренгиальной области зараженных животных. Титр нейтрализующих антител in vitro не всегда коррелировал с защитой животных.
Так как все полевые штаммы ВПБ содержат gE, а большинство аттенуированных штаммов, используемых в качестве живых вакцин, не содержат gE, появилась возможность использовать ИФА для дифференциации иммунных и инфицированных животных и тем самым способствовать оздоровлению хозяйств на основе вакцинопрофилактики. Указанная стратегия успешно реализуется в практических условиях в Западной Европе.
За индукцию синтеза ВН-антител герпесвируса типа I крупного рогатого скота (ГВ I) ответственны четыре гликопротеина оболочки. Главными из них являются g70 и g97. Эти гликопротеины содержат основные эпитопы нейтрализации вируса. Гемагглютинирующая активность ГВ крупного рогатого скота связана с мажорным g97, расположенным в шипиках вириона. Главные нейтрализующие эпитопы расположены на гемагглютинирующем домене, что свидетельствует о перспективности использования гемагглютинирующего антигена ГВ 1 крупного рогатого скота для приготовления субъединичной вакцины, 15381. ГВ1 крупного рогатого скота так же, как герпесвирус типа I кошек (ГВК1), агглютинирует эритроциты мышей, хотя гемагглютинирующие домены этих вирусов не идентичны.
В структуре gIV ГВ1 крупного рогатого скота идентифицированы три нейтрализующих домена. Домен I содержит два перекрывающихся эпитопа, домен III - четыре идентичных или перекрывающихся эпитопа, домен II - один эпитоп. Домен I имеет отношение к проникновению вируса в клетки-мишени. Прикрепление вируса к клеткам in vitro опосредовано его главными гликопротеинами, которые в этом отношении можно расположить в ряду gill > gl > gIV. Очищенный gIII полностью блокирует адсорбцию полевого штамма ГВ1 крупного рогатого скота.
В становление гуморального и клеточного иммунитетов свой вклад вносят гликопротеины оболочки gI gIII и особенно gIV. Максимальную защиту телят в экспериментальных условиях обеспечивал gIV, а также безвирусный лизат инфицированных клеток.
Нейтрализующие антитела к трем оболочечным гликопротеинам (gI, gIII и gIV) FBI крупного рогатого скота вводили телятам и определяли защитный эффект при экспериментальном заражении гомологичным вирусом. У привитых животных титры антител в носовом секрете и смывах с конъюнктивы были аналогичны таковым у реконвалесцентов. Несмотря на клинический эффект от введения антител, они не предотвращали размножение ГВ1 крупного рогатого скота в слизистой оболочке носа и глаз, а также не сокращали его экскрецию. Цитотоксические Т-лимфоциты крупного рогатого скота, индуцированные ГВ1 крупного рогатого скота, проявляют специфичность, то есть реагируют только на клетки, экспрессирующие гомологичный или родственный антиген. Мутант ГВ1 крупного рогатого скота с делецией гена gIII не теряет способности размножаться в культуре клеток. Его функциональная дефектность проявляется в замедленной адсорбции и репликации, а также в снижении выхода внеклеточного вируса. По аналогии с вирусом псевдобешенства идентифицированы и делетированы несущественные гены с целью получения аттенуированных «маркированных» вакцин.
Дата добавления: 2016-04-14; просмотров: 1111;