МЕХАНИЗМ ОБРАЗОВАНИЯ УСЛОВНЫХ РЕФЛЕКСОВ
Согласно И. П. Павлову (1903), временная связь образуется между корковым центров безусловного рефлекса и корковым центром анализатора, на рецепторы которого действует условный раздражитель, т. е. связь замыкается в коре большого мозга (рис. 13.4). В основе замыкания временной связи лежит процесс доминантного взаимодействия между возбужденными центрами. Импульсы, вызываемые индифферентным (условным) сигналом с любого участка кожи и других органов чувств (глаз, ухо), поступают в кору большого мозга и обеспечивают там образование очага возбуждения. Если после индифферентного сигнала подается пищевое подкрепление (подкормка), то возникает более мощный второй очаг возбуждения в коре больших полушарий, к которому направляет-
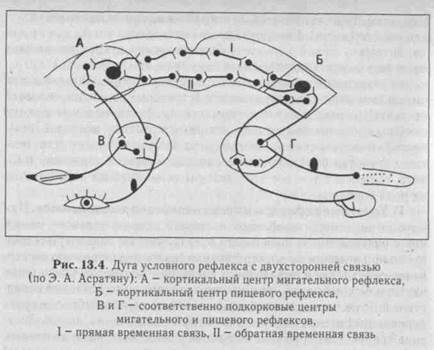
ся ранее возникшее и иррадиирующее по коре возбуждение. Неоднократное сочетание в опытах индифферентного (условного) сигнала и безусловного раздражителя (подкрепление) облегчает про-, хождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса - синапти-ческое облегчение (проторение пути). Образование временной связи в коре больших полушарий И. П. Павлов назвал замыканием новой условнорефлекторной дуги: теперь подача только условного сигнала приводит к возбуждению коркового центра безусловного рефлекса и возбуждает его, т. е. возникает рефлекс на условный раздражитель - условный рефлекс. Э. А. Асратян показал, что образование условных рефлексов с прямыми и обратными связями является общей закономерностью высшей нервной деятельности.
Следует отметить, что индифферентный сигнал не является вообще индифферентным, так как в начале его действия возникает реакция в виде ориентировочно-исследовательского рефлекса, который по мере повторения ослабляется.
На клеточном и молекулярном уровнях временная связь замкнется с помощью механизмов памяти.
ПАМЯТЬ
Память - способность организма приобретать, сохранять и воспроизводить в сознании информацию и опыт.
Биологическое значение памяти. Накопление, хранение и воспроизведение в сознании информации - общие свойства нейронных сетей. Невозможно переоценить биологическое значение этих процессов для адаптации индивидуального поведения к окружающей среде. Без способности к научению и памяти ни отдельная особь, ни вид в целом не могли бы выжить, поскольку оказались бы невозможными как планирование успешных действий, так и преднамеренное избегание ошибок. Соответственно, в последние десятилетия нейробиологи уделяли этим процессам много внимания. Было обнаружено, что запоминается лишь ничтожная доля общей осознаваемой сенсорной информации. С другой стороны, большинство накопленных сведений со временем, несомненно, забывается. Без таких механизмов отбора и забывания информации мы были бы буквально затоплены ее непрерывным потоком; результаты этого были бы такими же катастрофическими, как и отсутствие способности к научению и памяти.
По длительности хранения информации различные авторы выделяют несколько вариантов памяти, однако их продолжительность не согласуется в разных классификациях. Выделяют также сенсорную память (неточное название - «иконическая память»), которая соответствует длительности ощущений после прекращения действия сигнала, например звука, света.
Во-первых, «сенсорную память» необосновано называют особым видом памяти, так как нет воспроизведения в сознании информации или опыта (воспоминания), а есть продолжение ощущения после выключения раздражителя - как результат инерционности анализаторов, например, ощущение прикосновения, которого уже нет.
Во-вторых, все противоречия о длительности хранения информации можно исключить, положив в основу классификации не длительность хранения информации, а механизмы - электрофизиологические процессы, биохимические реакции и структурные изменения в нейронах и синапсах ЦНС. При этом идея длительности хранения информации также сохранится - кратковременная, промежуточная и долговременная память.
Кроме того, в раннем онтогенезе следует выделить память импринтинга(см. гл. 13.5). Механизмы импринтинга связаны с экспрессией в нейронах мозга специфических ранних генов - с-[оз и с-]'ип, функцией которых является перестройка работы генети-
ческого аппарата нервных клеток под влиянием запечатлеваемого воздействия. По механизму импринтинга запечатляется у взрослых животных действие жизненно значимых подкрепляющих факторов. По мере индивидуального развития животных механизм импринтинга все больше уступает место другим механизмам памяти.
А. Кратковременная память обеспечивает удержание и воспроизведение оперативной информации. Ее объем составляет примерно 7±2 единицы. Основным свойством данной разновидности памяти является ее непродолжительность. Хранение информации в кратковременной памяти длится секунды, минуты.
Механизм кратковременной памяти. Единой теории до сих пор не существует. Однако ясно, что запоминание любой информации начинается с электрофизиологических процессов в нейронных сетях головного мозга (возникновение ВПСП, ПД, выделение различных медиаторов в синаптическом аппарате мозга). Поэтому можно утверждать, что в основе механизма кратковременной памяти лежит импульсная активность нейронов и, в частности, циркуляция возбуждения по замкнутым нейронным цепям.
Большую роль для выяснения механизмов памяти сыграло применение в лечебных целях электрошока - сильного электрического воздействия на головной мозг. Как выяснилось, после электро-" шока, так же как и после механической травмы мозга в результате сотрясения, наркоза, наблюдается ретроградная амнезия, проявляющаяся в том, что человек полностью забывает о событиях, предшествовавших электрическому или механическому воздействию на мозг. По длительности ретроградной амнезии можно получить представление о времени консолидации памяти (лат. сопзоНаа.хю -упрочение, укрепление, объединение) - времени перехода кратковременной памяти в долговременную. Консолидация памяти начинается обычно через несколько минут, иногда через несколько десятков минут (в опытах на крысах, у которых после выработки условного рефлекса вызывали электрошок, приобретенный навык сохранялся, если электрошок проводился через 10 и более минут после начала обучения, при более раннем воздействии на мозг навык исчезал).
Наблюдения за развитием состояния ретроградной амнезии у человека привели к формулировке одной из первых гипотез о механизме кратковременной памяти - гипотезе реверберации (циркуляции) возбуждения по замкнутым цепочкам нейронов. Считают, что в замкнутых нейрональных цепочках циркуляция длится минутами, сохраняя информацию в виде последовательности импульсов, передающихся от нейрона к нейрону (см. рис. 5.8). Допус-
кают, что пока циркуляция продолжается, сохраняется нейрональ-ный след о воздействии того или иного раздражителя на организм в прошлом. С позиций гипотезы реверберации легко объясняется феномен ретроградной амнезии. Полагают, что развитие амнезии при травмах головного мозга, при электрошоке возникает вследствие грубого нарушения ритмической активности в цепочках взаимосвязанных нейронов.
Гипотеза реверберирующего возбуждения согласуется с нашим повседневным опытом, свидетельствующим о том, что для обучения необходима практика, т.е. неоднократное «пропускание» материала через сознание. Согласно морфологическим и электрофизиологическим данным, подобная реверберация по крайней мере возможна. Так, при выработке инструментальных условных рефлексов усвоение навыка сопровождается вполне определенными изменениями ЭЭГ (в частности, амплитуды вызванных потенциалов).
Результаты исследований подтверждают представление об электрофизиологическом происхождении кратковременной памяти (ВПСП, ПД, медиаторы), поэтому ее можно назвать электрофизиологической памятью.
Б. Промежуточная память - это процесс перевода кратковременной памяти в долговременную (консолидация памяти), который продолжается несколько часов. Следы кратковременной памяти становятся устойчивыми через 4 ч. Таким образом, длительность промежуточной памяти от нескольких минут до 4 ч. На данном этапе электрофизиологические процессы запускают с помощью вторых посредников биохимические реакции. Поэтому данный период переработки поступившей информации и запоминания ее следует назвать нейрохимической памятью.По длительности хранения информации ее следует назвать промежуточной памятью (между кратковременной и долговременной). В этот период экстремальные воздействия (наркоз, сотрясение мозга и др.) еще способны стереть память. Введение крысам блокатора синтеза ДНК - азидотимина - препятствует переходу промежуточной памяти в долговременную. Этот вид памяти изучен пока очень слабо.
В. Долговременная память. Ее основой являются структурные изменения в нейронах, длительность - часы - дни и на протяжении всей жизни при повторении информации, ее объем практически безграничен. Долговременная память устойчива к мозговым нарушениям. Она формируется с помощью кратковременной и промежуточной памяти, при этом важную роль играют синаптические процессы.
Важную роль в консолидации памяти играют нейропепти-ды. Показано, что пептиды могут находиться в пресинаптических терминалях в качестве сопутствующих медиаторов. Например, вместе с норадреналином часто выделяются нейропептид У, опиоидные пептиды, соматостатин. Дофамин часто выделяется окончаниями аксонов вместе с холецистокининином, энкефалином; ацетилхолин - с вазоактивным интестинальным пептидом, энкефалином, люлиберином; серотонин - с веществом Р, тиреолиберином, холецистокинином (И. П. Ашмарин). Доказано, что выделение пептидов в пресинаптических окончаниях зависит от частоты работы нейрона, при этом избыточное выделение пептида-спутника всегда наблюдается при усилении активности нейронов.
Роль синтеза белка. Активно разрабатывается гипотеза X. Хи-дена о белковой природе памяти, согласно которой процесс фиксации информации в нервной клетке находит отражение в синтезе белка, в молекулу которого вводится соответствующий следовой отпечаток изменений молекул РНК. При этом молекула белка становится чувствительной к специфическому узору импульсного потока, т. е. она узнает афферентный поток импульсов. Гипотеза о белковой природе долговременной памяти подтверждается экспериментально.
Так, если синтез белка, обычно начинающийся в мозгу животного во время сеанса выработки условного рефлекса и длящийся много часов, блокировать, долговременного научения не произойдет. Его подавление не влияет (по крайней мере, в опытах на животных) на кратковременную память. Это еще один важный аргумент в пользу того, что механизмы кратковременной и долговременной памяти различны. Однако процессы перевода информации из одной формы в другую остаются невыясненными. На основании изложенного все виды памяти по механизмам их формирования обосновано назвать: электрофизиологическая (кратковременная), нейрохимическая (промежуточная) и структурная (долговременная) память.
Гиппокамп выступает первым пунктом конвергенции условных и безусловных стимулов, он обеспечивает консолидацию памяти -перевод кратковременной памяти в долговременную.
Мозг работает как единое целое, но имеется и специализация -конкретный процесс научения затрагивает определенную популяцию нейронов и областей мозга. Так, височная кора участвует в запечатлении и хранении образной информации, патология височной коры ведет к потере долговременной памяти. При нарушении лобных долей отмечаются затруднения в организации действий, легкая отвлекаемость.
Дата добавления: 2016-04-11; просмотров: 992;