Типы клеточной организации.
1. Клетка – элементарная генетическая и структурно-функциональная биологическая система.
Клетка является элементарной, т.е. наименьшей, самостоятельной единицей строения, функционирования и развития всех живых организмов. Все живые организмы состоят из клеток (исключение составляют вирусы).
Клетка является генетической единицей, так как имеет ядро, содержащее ДНК, в которой записана информация о строении и функциях клетки. При делении клетки эта информация наследуется дочерними клетками.
Клетка является структурно-функциональная биологическая система: живая клетка включает в себя подсистемы – органеллы, органеллы выполняют определенные функции и в тоже время они все вместе обеспечивают жизнедеятельность клетки.
2. Клеточная теория. Современное состояние клеточной теории.
Началом изучения клетки можно считать 1665г., когда Р. Гук увидел в микроскоп на тонком срезе пробки мелкие ячейки, названные им клетками. В 30-е годы XIX века Р. Броун открыл ядро, что создало основу для сопоставления всех клеток.
В 1839г. Т. Шванн и М. Шлейден обобщили накопленный материал и сформулировали основное положение: все растительные и животные организм состоят из клеток, сходных по строению.
В 1858г. Р. Вирхов внёс очень важное дополнение в клеточную теорию – он доказал, что количество клеток увеличивается только одним способом – в результате деления. Таким образом, клетка происходит только от клетки.
Благодаря применению физических и химических методов исследования и новейших приборов, основные положения клеточной теории были развиты и углублены. Современная клеточная теория включает следующие положения:
1. клетка – основная единица строения и развития всех живых организмов, наименьшая единица живого;
2. клетки всех одноклеточных и многоклеточных организмов сходны (гомологичны) по своему строению, химическому составу, основным проявлениям жизнедеятельности и обмену веществ;
3. клетки размножаются делением, и каждая новая клетка образуется в результате деления исходной (материнской) клетки;
4. клетки многоклеточных организмов специализированы по выполняемым ими функциям и образуют ткани;
5. ткани образуют органы, которые тесно взаимосвязаны и подчинены нервным и гуморальным системам регуляции.
Вывод.
Клетка – живая элементарная открытая система, являющаяся основной структурно-функциональной единицей всех живых организмов, способная к самообновлению, саморегуляции и самовоспроизведению.
3. Клетка как открытая система. Организация потоков вещества, энергии и информации в клетке.
Клетка является открытой системой. Это значит, что в неё из внешней среды (если это организм человека, то из ЖКТ) поступают расщеплённые до мономеров питательные вещества, с заключенной в них энергией. Эти вещества используются в процессе жизнедеятельности клетки, а образующиеся при этом продукты метаболизма удаляются из него.
Поток вещества: человек съел яйцо → в ЖКТ оно переварилось → аминокислоты всосались в кровь и поступили в клетку → из них синтезируются видоспецифические белки, который обеспечивает жизнедеятельность клетки, → в процессе жизнедеятельности клетки белки изнашиваются, стареют, разрушаются → конечные продукты обмена белка (аммиак и вода) выводятся из клетки.
Поток энергии тесно связан с потоком вещества. Организм поглощает пищу (питание необходимо для обеспечения организма энергией и строительными материалами для репарационных и просто строительных целей) → часть пищи преобразуется в митохондриях в АТФ, а часть запасается в виде моносахаридов в печени, остальные углеводы разлагаются до Н2О и СО2.
Поток информации в клетке – в клетке постоянно происходит синтез белка. Информация о структуре белка записана в ДНК. В процессе транскрипции она переносится на и-РНК, а в процессе трансляции на полипептид.
Кроме ДНК и иРНК в реализации наследственной информации в клетке участвуют тРНК, рибосомы, ферменты активации аминокислот, АТФ.
4. Типы клеточной организации.
Все организмы, имеющие клеточное строение, делятся на две группы: прокариоты (про – до, карион – ядро), или предъядерные.
· эукариоты (эу – настоящие, карион – ядро), или ядерные.
Клетки прокариот (эубактерии и цианобактерии) имеют более простое строение:
1. нет организованного ядра, т.е. ядерное вещество не отделено от цитоплазмы собственной мембраной. Ядерное вещество представлено единственной хромосомой, состоящей из 1 молекулы ДНК, замкнутой в кольцо;
2. из органоидов присутствуют только многочисленные, но мелкие рибосомы;
3. функцию митохондрий у прокариот выполняют ферменты, лежащие непосредственно на плазматической мембране и образующие дыхательную цепь;
4. нет клеточного центра, следовательно, нет митоза (делятся амитозом);
5. не характерен циклоз (постоянное круговое движение цитоплазмы с органоидами), в то время как отсутствие циклоза для эукариот означает гибель клетки;
6. отсутствуют внутренние мембраны, делящие клетку на отсеки, в которых протекают противоположные процессы.
При всей простоте строения прокариоты – типичные клетки, способные вести независимое существование.
5. Организация эукариотической животной клетки.
· имеется оформленное ядро, в котором располагается молекула ДНК;
· имеется развитая система мембран и соответственно органоиды мембранного строения: ЭПС, аппарат Гольджи, лизосомы, митохондрии;
· имеется клеточный центр, поэтому размножаются митозом;
· могут передвигаться при помощи ложноножек (амеба);
· имеется циклоз;
· присуще явление компартментации – с помощью биологических мембран обеспечивается пространственное разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой или ее частью (пространство, отграниченное внутренней мембраной митохондрии)
6. Строение и функции оболочки животной эукариотической клетки.
Эукариотическая клетка отделена от внешней среды или соседних клеток плазматической мембраной, или плазмалеммой.
Среди многочисленных моделей мембран, наиболее универсальной оказалась так называемая "жидкостно-мозаичная" модель. Согласно ей основой мембраны является жидкостный билипидный слой, образованный строго ориентированными фосфолипидными молекулами. Двойной слой фосфолипидных молекул обращен друг к другу гидрофобными участками, а внешняя и внутренняя поверхности билипидного слоя образованы гидрофильными участками молекул. Белки, входящие в мембрану, не составляют сплошного слоя на внутренней и внешней поверхности билипидного слоя; они расположены мозаично и обладают способностью к перемещению в билипидном слое. Мембранные белки представлены тремя разновидностями:
· периферические белки располагаются на поверхности билипидного слоя;
· погружённые белки пронизывают всю толщу мембраны;
· полупогружённые белки погружены в мембрану лишь наполовину, выступая наружу с какой-то одной (внешней или внутренней) поверхности мембраны.
Из этой модели организации мембраны вытекает важное следствие, а именно: возможность горизонтального и отчасти вертикального смещения
погружённых и полупогружённых белковых молекул, то есть подвижность такой системы.
Пронизывающие белки участвуют в транспорте веществ.
Полупогружённые белки, обращённые внутрь, выполняют регуляторные ф-и.
Полупогружённые белки, обращённые наружу, «узнают» поверхность соседних клеток; благодаря им формируются ткани и органы.
На плазмалемме животных клеток находится гликокаликс – соединение белков и полисахаридов. Он непосредственно связывает клетку с внешней средой и служит для распознавания сигналов, поступающих из неё. Он же связывает клетки в ткани. Образуется гликокаликс благодаря жизнедеятельности самих клеток.
Функции плазматической мембраны:
· защитная или барьерная функция
· обеспечение контактов между клетками
· сигнальная (рецепторная) – на поверхности мембраны находятся рецепторы, которые воспринимают сигналы из внешней среды
· транспортная – регулирует транспорт в-в, т. к. обладает избирательной проницаемостью.
7. Трансмембранный транспорт веществ в клетке.
Существует 5 способов поступления веществ в клетку: диффузия, облегчённая диффузия (пассивный транспорт), осмос, активный транспорт, эндоцитоз. Два последних способа сопровождаются затратами энергии.
Диффузия – перемещение вещества из области большей концентрации в область меньшей концентрации без затраты энергии. Движущей силой диффузии является градиент (разность) концентрации. Диффузия будет действовать до тех пор, пока концентрация вещества в двух областях не выровняется. Так перемещаются кислород, углекислый газ, глюкоза, аминокислоты, жирные кислоты, этанол, мочевина.
Облегчённая диффузия или пассивный транспорт. В этом случае молекула специального белка-переносчика соединяется с переносимой молекулой на одной стороне мембраны и «перетягивает» её на другую сторону мембраны. Перенос веществ в этом случае осуществляется также по градиенту концентрации и без затраты энергии. Так перемещаются сахара, аминокислоты, нуклеотиды и ионы.
Осмос – перемещение молекул воды через полупроницаемую мембрану, вызванное разностью концентрации. Клетка, помещённая в чистую воду, насасывает её по градиенту своей концентрации. Клетка, помещённая в насыщенный раствор, отдаёт воду и сморщивается. Например, эритроциты в гипотоническом растворе набухают и лопаются, а в гипертоническом – сморщиваются.
Активный транспорт – перемещение веществ против их градиентов концентрации, т.е. из области меньшей концентрации в область большей концентрации. На это требуется энергия, источником её служит АТФ. Этот способ характерен только для ионов питательных веществ, следовательно, клетка обладает избирательной способностью по отношению к различным ионам.
Эндоцитоз – перемещение веществ с помощью выростов и выпячиваний плазмалеммы. Эндоцитоз делится на фагоцитоз и пиноцитоз. Оба этих процесса требуют затраты энергии.
Фагоцитоз – захват выростами плазмалеммы крупных частиц, диаметром 1мм и более и втягивание их в цитоплазму клетки. Здесь происходит их ферментативное расщепление. Клетки, осуществляющие фагоцитоз, называются фагоцитами (нейтрофилы и макрофаги).
Пиноцитоз – поглощение пузырьков жидкости. В месте соприкосновения с пузырьком плазмалемма образует впячивание в виде канальца, который заполняется жидкостью. Затем он отшнуровывается и попадает в цитоплазму. Пиноцитоз характерен для лейкоцитов, клеток зародыша и печени. Путём пиноцитоза поглощаются крупные молекулы и ионы, не способные проникать через поры плазмалеммы.
Из клетки во внешнюю среду вещества поступают с помощью экзоцитоза. Так из секреторных клеток, через их мембрану, выводятся пищеварительные ферменты или гормоны, а из пищеварительных вакуолей – непереваренные плотные частицы.
8.Цитоплазма: основное вещество, цитоскелет, органеллы.
Основное вещество цитоплазмы представлено гиалоплазмой. Это коллоидный раствор неорганических и органических веществ, особенно много в гиалоплазме белков.
Функции гиалоплазмы:
· соединение компонентов цитоплазмы в единое целое
· участие в транспорте веществ
· в гиалоплазме протекает гликолиз
· в гиалоплазме накапливается АТФ и включения.
Цитоскелет клетки представлен микротрубочками и микрофиламентами.
Каждая микротрубочка представляет собой полый цилиндр диаметром 20-30нм, образованный белком тубулином. Микротрубочи играют роль цитоскелета, т.к. пронизывают всю цитоплазму клетки. Кроме того, микротрубочки участвуют в создании клеточного центра и в транспорте веществ внутри клетки.
Микрофиламенты – это белковые нити толщиной около 4нм. Большинство из них образовано молекулами актинов, которых выявлено около 10 видов. Они могут группироваться в пучки, образующие опорные структуры цитоскелета.
Микротрубочки – трубчатые образования белковой природы различной длины с внешним диаметром 24 нм. Микротрубочки встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей.
Органеллы животной клетки: ЭПС, аппарат Гольджи, лизосомы, митохондрии, пластиды, рибосомы, клеточный центр.
Органоиды клетки делятся на органоиды общего назначения и специального назначения.
Органоиды специального назначения встречаются только в специализированных клетках и обеспечивают выполнение этими клетками специфических функций. К ним относятся миофибриллы мышечной клетки, ресничный эпителий дыхательных путей, ворсинки тонкого кишечника, жгутик сперматозоида.
Органоиды общего назначения присущи всем клеткам. К ним относятся эндоплазматическая сеть, лизосомы, митохондрии, рибосомы, комплекс Гольджи, клеточный центр, микротрубочки и микрофиламенты, а также пластиды (последние только у растений).
Эндоплазматическая сеть представлена сетью каналов и уплощённых цистерн, ограниченных одинарной мембраной. Она разветвляется по всему объёму цитоплазмы, что позволяет ей выполнять следующие функции:
· механическая – обеспечение постоянной формы клетки;
· увеличение площади внутренней поверхности клетки;
· транспортная – перенос веществ между органоидами клетки, органоидами и ядром, клеткой и внешней средой;
ЭПС подразделяется на два типа: шероховатую и гладкую. Шероховатая имеет на наружной поверхности многочисленные рибосомы, на которых синтезируется белок. Гладкая сеть состоит из каналов и цистерн меньшего сечения, чем в шероховатой ЭПС. Она выполняет следующие функции:
· синтез липидов, входящих в состав мембран;
· обезвреживание вредных продуктов метаболизма;
· синтез предшественников стероидных гормонов;
Лизосома. Это пузырёк диаметром 0,2 - 0,5 мкм, покрытый однослойной мембраной. Эта мембрана предохраняет структуры и вещества клетки от разрушающих действий ферментов лизосом. При нарушений её целостности ферменты выходят в цитоплазму клетки, и происходит автолиз – самопереваривание клетки. Ферменты лизосом способны расщеплять белки, нуклеиновые кислоты, полисахариды и липиды.
Функции лизосом:
1. осуществляют внутриклеточное пищеварение; лизосомы – миниатюрная пищеварительная система клетки;
2. удаляют отжившие органоиды клетки или личиночные органы. Так, хвост у головастика лягушек рассасывается под действием фермента лизосом – катепсина;
3. превращают вредные для клетки вещества в перевариваемые продукты;
4. участвуют в защите клетки то бактерий и вирусов (вирусы замуровываются в лизосоме).
Образуются лизосомы в комплексе Гольджи: сюда поступают синтезированные на рибосомах ферменты, здесь они окружаются мембраной и выводятся в цитоплазму. Это первичные (неактивные) лизосомы. Вторичные (активные) лизосомы образуются из первичных. Они подразделяются на фаголизосомы и аутолизосомы. Фаголизосомы переваривают материал, поступающий в клетку извне. Аутолизосомы разрушают собственные, изношенные структуры клетки. Вторичные лизосомы, в которых процесс переваривания завершён, называются остаточными тельцами. В них отсутствуют ферменты, и содержится непереваренный материал.
Митохондрия (1-5 мкм) – двумембранный органоид, выполняющий функцию внутриклеточной энергетической станции. Это округлые образования, ограниченные двумя мембранами – наружной и внутренней. Наружная мембрана гладкая, она регулирует как поступление веществ в митохондрию, так и выведение их. Внутренняя мембрана образует складки – кристы, обращённые внутрь митохондрии. Внутри митохондрии находится так называемый матрикс, содержащий различные ферменты, ионы Са2+ и Мg2+, а также ДНК, т-РНК, и-РНК и рибосомы (причём ДНК и рибосомы у митохондрий похожи на таковые бактерий).
Благодаря наличию собственной ДНК (1 молекула кольцевой формы), митохондрии могут размножаться независимо от деления клетки. Происходит это путём перешнуровки исходной митохондрии. Предварительно у них удваивается количество ДНК. Благодаря содержанию т-РНК, и-РНК и рибосом, митохондрии они могут синтезировать собственный белок.
Кроме того, митохондрии играют определённую роль в передаче признаков по наследству (цитоплазматическая наследственность).
На кристах митохондрии происходят окислительно-восстановительные процессы, сопровождающиеся выделением энергии. Она используется на образование фосфатных связей в АТФ. Накопление АТФ делает митохондрии своеобразными аккумуляторами энергии клетки, которая расходуется на процессы жизнедеятельности клетки по мере надобности. Из-за интенсивной работы митохондрии имеют малую продолжительность жизни, например митохондрии клеток печени живут всего 10 дней.
Аппарат Гольджи представляет собой систему диктиосом числом от нескольких десятков до нескольких сотен и даже тысяч на клетку. Каждая диктиосома образована стопкой из 3-12 крупных цистерн, похожих на блюдца. От цистерн отходят во все стороны трубочки и пузырьки, имеющие мембранное строение. Трубочки соединяют отдельные цистерны соседних стопок, так образуется их единая сеть. Пузырьки участвуют в образовании первичных лизосом. В разных типах клеток аппарат Гольджи занимает строго определённое положение, вблизи ядра.
Функции аппарата Гольджи разнообразны:
1. образование первичных лизосом, которые поступают затем в цитоплазму;
2. упаковка белков, поступающих из ЭПС, для последующего экспорта из клетки;
3. синтез структурных компонентов клетки, например, коллагеновых нитей;
4. синтез жиров и полисахаридов, входящих в состав мембран клетки;
Рибосома (0,02-0,03 мкм) – не мембранный органоид, осуществляющий биосинтез белка. Рибосома состоит из двух неравных субъединиц – большой и малой. Обе субъединицы образуются в ядрышках, но объединяются они в рибосому только в момент присоединения к и-РНК. Этот процесс происходит с помощью ионов Мg2+.
В каждой клетке содержится от десятков тысяч до миллионов рибосом. Часть их находится в свободном состоянии, но большинство рибосом прикреплено к мембранам ЭПС. Первые синтезируют белки, используемые для нужд клетки, вторые синтезируют белки "на экспорт". Они по каналам ЭПС поступают в комплекс Гольджи, пакуются в мембраны, а затем выводятся из клетки. Скорость работы рибосом поразительна – одна белковая молекула средних размеров синтезируется за одну минуту. Это позволяет непрерывно обновлять белки организма, изнашивающиеся в процессе его жизнедеятельности. Так, белки печени человека обновляются за 7 суток.
Клеточный центр – не мембранный органоид, в котором из белка тубулина образуются микротрубочки. Клеточный центр состоит из двух центриолей, расположенных перпендикулярно друг к другу. Каждая центриоль – это цистерна, состоящая из 9 строенных микротрубочек. Микротрубочки соединены между собой системой связок, а снаружи одеты белковым чехлом. Перед делением клетки центриоли удваиваются. Во время митоза центриоли определяют местоположения полюсов веретена деления. Причём положение центриолей в делящейся клетке определяет центры новых клеток. Здесь будет располагаться ядро, т.к. клеточный центр всегда располагается вблизи ядра.
9. Включения
Это непостоянный компонент цитоплазмы. Наличие их и количество зависит от интенсивности обмена веществ и состояния организма. Они делятся на три группы:
1. запасной питательный материал (гликоген, жир, крахмал);
2. вещества, подлежащие выведению из клетки (ферменты, гормоны);
3. балластные вещества (пигменты, соли щавелевой кислоты). Они более характерны для растительных клеток, т.к. у растений нет систем, аналогичной выделительной системе животных.
ЛЕКЦИЯ 2Ядро клетки. Наследственный аппарат клеток человека.
1. Строение и функции ядра.
Ядро находится либо в центре клетки, либо смещено на периферию. Ядро эукариотической клетки имеет собственную мембрану, отграничивающую его от цитоплазмы. Мембрана имеет 2 слоя, между ними находится околоядерное пространство, связанное с ЭПС.
Ядерная мембрана имеет отверстия – поры. Но они не сквозные, а заполнены специальными белками. Через поры из ядра в цитоплазму выходят молекулы РНК, а навстречу им в ядро передвигаются белки. Сама же мембрана ядерной оболочки обеспечивает прохождение низкомолекулярных соединений в обоих направлениях. Внутренняя мембрана ядерной оболочки имеет белковую подстилку, к которой крепятся хромосомы. Это обеспечивает их упорядоченное расположение.
Функции ядерной оболочки: защитная, регуляция транспорта веществ и органелл
Под мембраной находится ядерный сок – кариоплазма. В ней находятся одно или несколько ядрышек, значительное количество РНК и ДНК, различные белки, в т.ч. большинство ферментов ядра, а также свободные нуклеотиды, аминокислоты, промежуточные продукты метаболизма. Кариоплазма осуществляет взаимосвязь всех ядерных структур.
Ядрышки – это округлые, сильно уплотнённые, не ограниченные мембраной участки клеточного ядра диаметром 1-2 мкм и больше. Форма, размеры и количество ядрышек зависят от функционального состояния ядра: чем крупнее ядрышко, тем выше его активность. В ядре их может содержаться от 1 до 10, а в ядрах дрожжей они отсутствуют.
Во время деления ядра ядрышки разрушаются. В конце деления они вновь формируются вокруг определённых участков хромосомы (ядрышковых организаторов), расположенных в области вторичной перетяжки хромосомы. Функция ядрышек состоит в синтезе
р-РНК и сборки субъединиц рибосом из белка и р-РНК.
2. Наследственный аппарат клеток. Химическая и структурная организация хромосом.
Основное вещество ядра – хроматин. Он состоит из ДНК (40%), основных белков, или гистонов (40%) и кислых белков (20%). Перед митозом хроматин уплотняется за счет спирализации ДНК и приобретает определённую форму. Теперь он называется хромосомой. Хромосома – структурное образование, хроматин – химический эквивалент хромосом.
Важную роль в структурной организации хроматина и хромосом играют белки гистоны. По химическим свойствам это щелочные (основные) белки, в их состав в большом количестве входят аминокислоты аргинин и лизин, эти аминокислоты имеют 2 аминогруппы и 1 карбоксильную группу. Белки гистоны несут (+) заряд, а ДНК (–) за счет остатка фосфорной кислоты, поэтому имеет место взаимодействие белков гистонов и ДНК. Выделяют 5 классов белков гистонов: H1, H2A, H2B, H3, H4.
Структурная организация хромосом достаточно сложная.
В интерфазном ядре принято выделять 3 уровня структурной организации хромосом:
1) образование нуклеосом. Белки гистоны H2A, H2B, H3, H4, (по 2 молекулы каждого) образуют основу нуклеосомы (остов катушки), вокруг этой основы ДНК делает 2 витка. Белок гистон H1 связывает соседние нуклеосомы, образуется нуклеосомная нить. Длина ДНК уменьшается в 7 раз.
2) образование фибрилл или спиралеподобной структуры. Белок гистон H1 скрепляет витки спирали. Длина ДНК на этом уровне уменьшается в 6 раз.
3 уровень. Образование петель. ДНК человека образует до 2 тыс. петель. Длина ДНК уменьшается в 25 раз.
Таким образом, в интерфазных хромосомах длина ДНК за счёт спирализации уменьшается ≈ в 1000 раз. Во время митоза происходит дальнейшая спирализация хромосом. Длина ДНК уменьшается в десять тысяч раз. Вероятно смысл компактизации хромосом, заключается в том, чтобы при митозе хромосомы расходились точно к полюсам и не мешали друг другу.
Хромосомы максимально спирализованы в метафазу митоза и их можно увидеть в световой микроскоп.
Каждая хромосома состоит из двух хроматид, которые соединяются с помощью первичной перетяжки (центромеры).
Иногда на одном из плеч хромосомы может находиться вторичная перетяжка, которая отделяет спутник. Такие хромосомы называются спутниковыми, у человека это хромосомы 13,14,15,21,22 пар. Область вторичной перетяжки называется ядрышковым организатором, т.к. здесь образуются ядрышки.
В зависимости о положения первичной перетяжки выделяют следующие виды хромосом:
1. равноплечие (метацентрические) – центромера делит хромосомы на два равных плеча
2. слабо неравноплечие (субметацентрические) – центромера делит хромосомы на два слабо неравных плеча
3. резко неравноплечие (акроцентрические) – центромера делит хромосомы на два резко неравных плеча
4. одноплечие (телоцентрические) – центромера располагается на самом конце хромосомы.
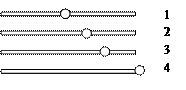 |
У человека нет телоцентрических хромосом.
3. Характеристика ДНК. Молекулярное строение гена у прокариот и эукариот.
Роль хранителя наследственной информации у всех организмов принадлежит ДНК. Эта кислота была открыта в 1869г. Ф. Мишером в ядрах лейкоцитов, но строение её было выяснено только в 1953г. Дж. Уотсоном и Ф. Криком. В своих исследованиях эти учёные опирались на данные рентгеноструктурного анализа молекулы ДНК и на установленное Э. Чаргаффом правило: в молекуле ДНК число пуриновых оснований строго соответствует числу пиримидиновых оснований.
ДНК – полимер, состоящий из десятков (или сотен) миллионов мономеров – дезоксирибонуклеотидов. В состав каждого дезоксирибонуклеотида входит азотистое основание, углевод (дезоксирибоза) и остаток фосфорной кислоты. Нуклеотиды ДНК отличаются друг от друга основаниями. Различают пуриновые основания: аденин (А) и гуанин (Г) и пиримидиновые основания: цитозин (Ц) и тимин (Т). Нуклеотиды ДНК называются соответственно: адениловый, гуаниловый, цитидиловый, тимидиловый.
Нуклеотиды ДНК соединены последовательно в цепочку за счет фосфодиэфирных мостиков, образующихся между углеводом одного нуклеотида и остатком фосфорной кислоты соседнего. Иначе говоря, остаток фосфорной кислоты связывает углеводы соседних нуклеотидов.
ДНК – это две правозакрученные цепи, основания которых обращены внутрь спирали и образуют пары таким образом, что (А) одной цепи всегда находится против (Т) другой цепи, а (Г) – против (Ц). Между этими парами оснований образуются водородные связи: две между А и Т и три между Г и Ц. В каждом сочетании оба нуклеотида как бы дополняют друг друга, они комплементарны. Комплементарность – взаимное соответствие в химическом строении молекул, обеспечивающее их взаимодействие. Комплементарные структуры подходят друг к другу как ключ к замку.
На один виток спирали ДНК приходится 10 нуклеотидных остатков. Т.к. расстояние между соседними нуклеотидами = 0,34 нм, шаг спирали составляет 3,4 нм.
Диаметр спирали – около 2 нм. Длина спирали может измеряться в метрах.
В процессе эволюции количество ДНК у эукариот возросло в 1000 раз, а количество генов увеличилось в 50 – 100 раз. Это говорит о том, что не вся ДНК в эукариотической клетке несет смысловую нагрузку, т.е. имеет место явление избыточности ДНК. В эукариотической клетке 99% ДНК избыточна, и только 1% несет смысловую нагрузку.
Геном эукариотической клетки устроен сложно, в нем выделяют 3 класса последовательностей нуклеотидов в ДНК.
1) многократно-повторяющиеся последовательности нуклеотидов, они составляют 15% ДНК. Эти последовательности содержат до сотни нуклеотидов и повторяются в геноме десятки тысяч, сотни тысяч и до миллиона раз (105-106 копий).
Предполагаемая роль многократно повторяющихся последовательностей:
а) узнавание гомологичных участков хромосом во время кроссинговера,
б) разделение структурных и регуляторных генов в кодирующих участках ДНК,
в) регуляция функции генов.
2) умеренно повторяющиеся последовательности нуклеотидов составляют от 10 до 50% ДНК (в геноме у человека 10%). Содержат тысячи нуклеотидов и повторяются в геноме десятки раз и десятки тысяч раз (102-104 копий).
Эти последовательности содержат гены, которые отвечают за синтез тРНК, рРНК, и белки гистоны. Особенностью этих последовательностей является наличие в них 2 участков: кодирующих информацию (гены) и не кодирующих (спейсеры).
3) уникальные гены, 75% ДНК имеют от 1 до 10 копий. Кодируют все белки организма (структурные и регуляторные), кроме белков гистонов.
Для уникальных генов характерно экзонно-интронное строение: кодирующий участок - экзон чередуется с не кодирующим участком - интроном.
4. Классификация хромосом.
1960г. Первая классификация хромосом в Денвере. Хромосомы стали нумеровать от первой пары до 23 пары.
В 1960 году Патау предложил разделить все хромосомы на 7 групп.
Группа Номера хромосом Виды хромосом
I (А) 1, 2, 3 большие метацентрические
II (В) 4, 5 большие субметацентрические
III (С) 6 – 12 средние субметацентрические
IV (D) 13, 14, 15 акроцентрические, спутничные
V (E) 16, 17, 18 малые субметацентрические
VI (F) 19, 20. малые метацентрические.
VII (G) 21, 22 акроцентрические спутничные
Отдельно выносят «Х» хр. и «У» хромосомы. «Х» хромосома – это средняя субметацентрическая хромосома. «У» хромосома – это акроцентрическая, но не спутничная.
5. Гетеро – и эухроматин.
В интерфазных хромосомах выделяют более спирализованные участки и менее спирализованные участки. Более спирализованные участки хорошо окрашиваются красителями и выглядят в световой микроскоп в виде глыбок – эти участки называются гетерохроматин, в этих участках нет функционирующих генов. Выделяют 2 вида гетерохроматина: структурный и факультативный.
Структурный гетерохроматин находится, как правило, на концах хромосомы и в области центромеры. В этих участках находятся многократно повторяющиеся последовательности ДНК, которые выполняют, как правило, структурную роль. Структурный гетерохроматин не содержит структурных генов и постоянно находится в спирализованном состоянии. Он составляет 16% генома.
Факультативный гетерохроматин. В клетках ♀ организма в норме находятся 2 «Х» хромосомы, но 1 «Х» хромосома находится в неактивном состоянии в виде гетерохроматина. В световой микроскоп в интерфазном ядре такая хромосома выглядит в виде плотного округлого тельца и называется она половой хроматин (или тельце Барра). Всегда в любом кариотипе функционально активна только одна «Х» хромосома, а остальные «Х» хромосомы переходят в состояние гетерохроматина (инактивируются на 16 день после оплодотворения).
Эухроматин – это менее спирализованные участки хромосом, которые плохо окрашиваются красителями. В этих участках находятся функционирующие гены. Меньшая спирализация этих участков позволяет быстрее списывать с них наследственную информацию.
6. Наследственный аппарат клеток человека. Кариотип человека, характеристика кариотипа в норме.
Кариотип – это хромосомный набор характерный для данного вида организмов и характеризующийся определенным числом и строением хромосом. Или совокупность данных о числе, форме и размерах хромосом называется – кариотип.
В кариотипе человека 23 пары хромосом. 22 пары хромосом одинаковы у мужчин и женщин, они называются аутосомами. Одна пара хромосом – половые хромосомы, у мужчин это хромосома Х и хромосома У, у женщин это две Х хромосомы.
Хромосомы изучают на стадии метафазы митоза, в этот период они максимально спирализованы и хорошо видны в световой микроскоп.
Метафазная пластинка. Для исследований у человека берут лейкоциты крови, а затем лейкоциты обрабатывают специальными веществами, которые запускают митоз (митогенами). Через 48 часов клетки будут на стадии митоза, и деление останавливают с помощью веществ, которые разрушают нити веретена деления (колхицин), затем хромосомы окрашивают и микроскопируют.
Число хромосом в гамете, свойственное данному виду, называется гаплоидным (1n), а число хромосом, свойственное зиготе и соматическим клеткам, диплоидным (2n). Рисунки или снимки хромосом, которые располагаются в порядке убывания их размера, носят названия кариограмм или идиограмм.
ЛЕКЦИЯ 3 Временная организация клеток. Митотический цикл. Размножение.
1. Закономерность существования клетки во времени. Клеточный цикл, его периодизация.
Все клетки имеют определенный жизненный цикл, это период с момента появления клетки из материнской, до собственного деления или гибели.
В жизненном цикле делящихся клеток выделяют: митотический цикл, а также периоды выполнения клеткой специальной функции и период покоя. В период покоя клетка “решает” по какому пути она пойдет: по пути специализации или по пути деления. Все определяет состояние макроорганизма в каждый данный момент. Биохимическая основа этого “решает” связана с накоплением тригерного (пускового) белка.
2. Митотический цикл, фазы авторепродукции и распределения генетического материала.
Митотический цикл – это совокупность процессов, которые происходят в клетке при подготовке к делению и в процессе самого деления. В митотическом цикле выделяют: интерфазу (фазу авторепродукции) и митоз (фазу распределения генетического материала).
Интерфаза – это период подготовки клетки к делению, она составляет до 90% времени митотического цикла.
Интерфаза состоит из 3х периодов.
· период G1 – постмитотический период или пресинтетический. Он занимает 30 – 40% времени интерфазы.
· период S – синтетический период. Он занимает 30% времени интерфазы.
· период G2 – постсинтетический или премитотический и составляет 20 – 30% времени интерфазы.
Период G1 (2n2c 2n4c) в клетке синтезируются все виды РНК, белки, достраиваются необходимые органоиды, т.е. клетка увеличивается в объеме. Также накапливаются предшественники нуклеотидов, из которых будет синтезироваться ДНК.
В этот период выделяют особый момент, который называется точкой рестрикции. Пройдя точку рестрикции, клетка обязательно пойдет по пути деления. Предполагают, что в этот момент в клетке накапливается особый пусковой (тригерный) белок который запускает удвоение центриолей, затем удвоение ДНК, и деление клетки.
В конце G1 периода начинается удвоение центриолей клеточного центра.
Период S (2n2c → 2n4c) продолжается синтез РНК, белков. Самое важное событие – это удвоение ДНК (репликация ДНК), это сложный ферментативный процесс, протекающий в ядре клетки, и требует расхода энергии АТФ.
Синтез новых цепей ДНК идет из свободных нуклеотидов присутствующих в ядре и основан на принципе комплементарности.
В репликации участвует много ферментов: геликаза, ДНК полимераза, лигазы.
Геликаза разрывает водородные связи между 2мя цепями ДНК в определенных участках. Такие участки называются репликационные вилки или репликоны (в каждой хромосоме может быть до 100 репликонов).
ДНК полимераза синтезирует новые цепи ДНК. 2 цепи ДНК антипаралельны, а фермент ДНК полимераза всегда двигается только в направлении 5'→3'. Одна новая цепь синтезируется быстрее и называется лидирующей или ведущей. Другая новая цепь ДНК синтезируется в виде отдельных фрагментов и называется отстающей. Затем эти фрагменты (фрагменты Оказаки) сшиваются ферментом лигазой.
В результате репликации из одной молекулы ДНК образуются 2 молекулы, в каждой молекуле одна цепь старая, другая – новая. Поэтому способ называется полуконсервативным. В синтетический период происходит также синтез белков гистонов. В результате каждая хромосома состоит из двух сестринских хроматид.
Период G2 (2n4c) продолжается синтез всех видов РНК, белков, АТФ, накапливается белок тубулин, заканчивается удвоения центриолей клеточного центра.
Митоз (кариокинез, или фаза распределения генетического материала) – непрямое деление клетки.
Профаза (2n4c).
Клетка округляется, увеличивается вязкость цитоплазмы, центриоли расходятся к разным полюсам клетки, образуется веретено деления, основу его составляют микротрубочки, состоящие из белка тубулина. Нити веретена деления (микротрубочки) которые идут от 1одного полюса клетки к другому полюсу называются непрерывными.
Начинается спирализация хромосом, разрушаются и исчезают ядрышки.
Прометафаза (2n4c).
Разрушается ядерная оболочка и содержимое ядра перемешивается с цитоплазмой. В области центромеры с обеих сторон формируются особые структуры – кинетохоры, это слоистые структуры, состоящие из белков. От кинетохоров отходят кинетохорные или прерывистые нити веретена деления. Кинетохорные нити скользят вдоль непрерывных нитей веретена деления, что вызывает движение хромосом. Хромосомы движутся к центру клетки, к экватору клетки, продолжается спирализация хромосом.
Метафаза (2n4c).
Хромосомы максимально спирализованы и располагаются по экватору клетки, образуя метафазную пластинку. Каждая хромосома состоит из 2х сестринских хроматид соединенных центромерой.
Анафаза (2n4c → 4n4c).
Начинается с момента разделения центромер, сестринские хроматиды или дочерние хромосомы расходятся к разным полюсам клетки. Для движения хромосом необходимы нити веретена деления и энергия АТФ.
Телофаза (4n4c → 2n2c + 2n2c).
Происходит деспирализация хромосом, они раскручиваются, образуются ядрышки, ядерная оболочка и, происходит деление цитоплазмы (цитокинез). Из одной материнской клетки образуются 2 дочерние клетки.
Митоз протекает с большой точностью: может возникнуть одна ошибка на 100.000 делений.
Биологическое значение митотического цикла состоит в обеспечении преемственности хромосом в ряду клеточных поколений, образовании клеток, равноценных по объему и содержанию наследственной информации.
Изменение наследственного материала в митотическом цикле.
Фаза митоза Генетическая формула
Профаза 2n4c
Прометафаза 2n4c
Метафаза 2n4c
Анафаза 2n4c → 2n2c (на полюсах клетки)
Телофаза 2n2c + 2n2c (в дочерних клетках)
n – количество хромосом, с – количество молекул ДНК.
3. Механизмы регуляции митотической активности.
В многоклеточном организме деление клеток или пролиферация строго регулируется. Выделяют 4 уровня регуляции пролиферации.
· Внутриклеточный уровень. В клетке есть вещества регулирующие пролиферацию: ионы Са2+, циклические нуклеотиды ц АМФ, ц ГМФ.
· Внутритканевой уровень. На этом уровне важную роль играют кейлоны – это пептиды, они подавляют деление клеток. Противоположным действием обладают антикейлоны.
· Межтканевой уровень. Важную роль играют факторы лимфоцитов – лимфокины. Есть активаторы и ингибиторы.
· Организменный уровень. Важную роль играют гормоны, а также нейромедиаторы, нейросекреты и белки сыворотки (плазмы) крови.
4. Проблемы клеточной пролиферации в медицине.
Избыточная пролиферация клеток приводит к развитию опухолей. Недостаточная пролиферация клеток приводит к атрофии тканей (органа), или долгом заживлении раны. В любом случае речь идет об отклонениях в регуляции пролиферации клеток.
5. Эндомитоз и политения, их значение. Прямое деление клетки – амитоз.
В ходе эволюции, на основе митотического цикла сформировались процессы, которые приводят к увеличению количества наследственного материала в клетке. А это приводит к увеличению количества генов и к повышению активности обменных процессов в клетке. К таким процессам относят эндомитоз и политению.
Эндомитоз – удвоение ДНК клетки сопровождающееся кратным увеличением количества хромосом (4n4c). Механизм эндомитоза: в интерфазу происходит репликацией ДНК, а цитоплазма не делится.
Политения заключается в кратном увеличении содержания ДНК в хромосомах при сохранении их диплоидного количества (2n4c).
Эндомитоз и политения приводят к образованию полиплоидных клеток, отличающихся кратным увеличением объема наследственного материала. Для млекопитающих полиплоидизация в норме не характерна (кроме клеток печени).
Амитоз – прямое деление клетки. Амитоз характерен для прокариот, у многоклеточных организмов описан для стареющих и патологически измененных клеток. При амитозе интерфазное ядро делится путем перетяжки, равномерное распределение наследственного материала не обеспечивается. Нередко ядро делится без последующего разделения цитоплазмы, и образуются двухъядерные клетки. Клетка, претерпевшая амитоз, в дальнейшем не способна вступать в нормальный митотический цикл. Поэтому амитоз встречается, как правило, в клетках и тканях, обреченных на гибель, например в клетках зародышевых оболочек млекопитающих, в клетках опухолей.
6. Размножение – универсальное свойство живого, обеспечивающее материальную непрерывность в ряду поколений.
Способность к размножению является неотъемлемым свойством живых существ. С его помощью сохраняются во времени биологические виды и жизнь как таковая. Биологическая роль размножения состоит в том, что оно обеспечивает смену поколений. Различия, закономерно проявляющиеся в фенотипах особей разных поколений, делают возможным естественный отбор и, следовательно, эволюцию жизни. Последнее связано с тем, что при размножении осуществляется передача в ряду поколений генетического материала (ДНК), т.е. определенной, специфичной для данного вида биологической информации.
7. Классификация способов размножения.
Существует два способа размножения организмов – бесполое и половое.
Бесполое размножение осуществляется в следующих формах:
1. Деление надвое (простейшие).
2. Множественное деление (малярийный плазмодий) – сначала многократно делится ядро, а затем вокруг каждой частицы ядра образуется участок цитоплазмы.
3. Почкование (кишечнополостные).
4. Фрагментация – распад организма на части, которые превращаются в полноценные организмы (иглокожие, плоские черви).
5. Вегетативное размножение осуществляется за счет группы соматических клеток растения (листья, корни, побеги и их видоизменения).
6. Спорообразование. Спора – специализированная клетка, имеющая приспособления для перенесения неблагоприятных условий.
Половое размножение осуществляется с помощью специализированных клеток (гамет): яйцеклеток и сперматозоидов.
8. Сущность различных форм размножения и их эволюционная характеристика.
Для всех способов бесполого размножения характерна одна черта – источником наследственной информации служит одна клетка или группа клеток одного родителя. Это ведёт к образованию копии материнского организма. Дочерние особи генетически однообразны, их фенотипы почти одинаковы; поэтому в такой группе особей естественный отбор мало эффективен. Преимущество бесполого размножения состоит в простоте и эффективности: не нужно находить партнера; потомство может оставить любая особь и в любом месте; удачные мутации не теряются, а воспроизводятся в каждом поколении.
При переходе к половому размножению эти преимущества утрачиваются. Однако в результате оплодотворения происходит комбинирование признаков мужских и женских организмов, чем обеспечивается генетическое разнообразие потомства. Возникает новый неисчерпаемый источник наследственной изменчивости, который делает естественный отбор эффективным. Появление полового размножения – важное эволюционное событие. Оно придаёт популяции пластичность, что особенно необходимо в изменчивой среде.
9. Половой диморфизм: генетический, морфофизиологический, эндокринный и поведенческий аспекты.
Разделение половых клеток на яйцеклетки сперматозоиды, половых желез на яичники и семенники, а особей на самок и самцов – называется половым диморфизмом.
Генетический аспект: в основе полового диморфизма лежат различия в хромосомном наборе мужских и женских и особей.
Морфофизиологичекий аспект: в связи с различием в хромосомном наборе мужских и женских и особей организмы имеют свое морфологическое строение, влияющее на физиологические функции и процессы.
Эндокринный аспект: мужские (тестостерон) и женские (эстрогены) половые гормоны оказывают различное воздействие на человека, что приводит к появлению вторичных половых признаков.
Поведенческий аспект: наличие мужских и женских вторичных половых признаков и интенсивная выработка половых гормонов в период полового созревания обуславливает различное поведение самцов и самок.
10. Партеногенез.
Партеногенез – это разновидность полового размножения, когда развитие организма происходит из неоплодотворенной яйцеклетки.
Партеногенез подразделяется естественный и искусственный. В свою очередь естественный партеногенез делится на облигатный (обязательный) и факультативный (необязательный).
При облигатном партеногенезе яйца способны только к партеногенетическому развитию. Такой партеногенез наблюдается у некоторых популяций скальных ящериц.
При факультативном партеногенезе яйца могут развиваться и посредством партеногенеза и в результате оплодотворения. Такой партеногенез наблюдается у пчел, когда трутни развиваются из неоплодотворенных яиц, а матка и рабочие пчёлы – из оплодотворённых яиц.
Искусственный партеногенез вызывают действием на яйца различных раздражителей: уколом иглой, обработкой растворами сильных кислот, резким охлаждением и особенно нагревом.
С помощью искусственного партеногенеза обычно удается получать лишь начальные стадии развития организма; полный партеногенез достигается редко, хотя известны случаи полного партеногенеза даже у позвоночных животных (лягушка, кролик).
Способ массового получения полного партеногенеза, разработанный (1936) для тутового шелкопряда Б.Л. Астауровым, основан на точно дозированном кратковременном прогреве (до 46ºС в течение 18 мин) извлеченных из самки неоплодотворенных яиц. Этот способ дает возможность получать у тутового шелкопряда особи только женского пола, наследственно идентичные с исходной самкой и между собой. Таких особей можно потом размножать посредством партеногенеза неограниченно долго.
11. Биологические аспекты репродукции человека.
Человек – раздельнополое млекопитающее. Имеет разделение половых клеток на яйцеклетки и сперматозоиды, половых желез на яичники и семенники, а также различия в хромосомном наборе ♀ и ♂ особей. Характерна медленная смена поколений, низкая плодовитость (обычно 1–2 ребенка, не сравнить с дрозофилой, или тараканом).
ЛЕКЦИЯ 4 Половые клетки. Гаметогенез. Мейоз.
1. Морфофизиологическая характеристика яйцеклеток. Типы яйцеклеток.
Яйцеклетка – это крупная, округлая, неподвижная клетка. Размеры варьируют, диаметр у млекопитающих 100 – 200 мкм (микрометров). Это типичная эукариотическая клетка. В ней выделяют: оболочку, цитоплазму и гаплоидное ядро.
В цитоплазме находятся органоиды и включения в виде желтка. В желтке много белка, а также находится жироподобное вещество лецитин. В зависимости от количества и распределения желтка по цитоплазме выделяют следующие типы яйцеклетки.
1. Изолецитальные – желтка мало и он равномерно распределен по цитоплазма. Ядро в них располагается ближе к центру (черви, моллюски, ланцетник, млекопитающие).
2. Телолецитальные – желтка много и он находится на вегетативном (растущем) полюсе клетки, а ядро будет на противоположном анимальном полюсе (рыбы, амфибии, птицы)
а) умеренно телолецитальные яйцеклетки содержат среднее количество желтка (осетровые рыбы, земноводные).
б) резко телолецитальные яйцеклетки содержат очень много желтка, занимающего почти весь объем клетки (некоторых рыбы, пресмыкающиеся, птицы, яйцекладущие млекопитающие).
3. Центролецитальные – желтка много и он находится в центре клетки (насекомые).
Все яйцеклетки имеют плазматическую мембрану. Яйцеклетки многих животных кроме плазматической мембраны имеют дополнительные защитные оболочки.
Первичная оболочка, или желточная. Она образуется в результате жизнедеятельности самой яйцеклетки. У млекопитающих её называют блестящей оболочкой или зона пеллюцида. Zona pellucida
Вторичная оболочка (хорион у млекопитающих) образуется в результате жизнедеятельности фолликулярных клеток, которые окружают яйцеклетку в яичнике.
Третичная оболочка образуется при прохождении яйцеклетка по яйцеводу. В яйцеклетке птиц это белковая оболочка, подскорлуповая, и скорлуповая оболочки
2. Морфофизиологическая характеристика сперматозоидов.
Сперматозоид человека (50мкм), состоит из головки, шейки, средней части и хвостика. Головка содержит ядро с ДНК, передняя часть головки содержит акросому – структуру, выделяющую ферменты для растворения оболочки яйцеклетки. Это видоизменённый комплекс Гольджи, содержащий лизосому. В шейке содержатся две центриоли: одна участвует в образовании веретена деления, а другая – оси хвоста. Средняя часть содержит митохондрии, обеспечивающие сперматозоид энергией во время движения. Хвостик – органоид движения.
3. Процесс образования половых клеток (гаметогенез).
Гаметогенез – процесс образования половых клеток.
Он делится на сперматогенез – развитие мужских половых клеток – и овогенез – развитие женских половых клеток. В гаметогенезе выделяют 5 периодов: обособление, размножение, рост, созревание и формирование.
1. Обособление – первичные половые клетки обособляются от соматических клеток. Однако, они содержат диплоидный набор хромосом, их генетическая формула 2n2c.
2. Размножение – первичные половые клетки (сперматогонии или овогонии) делятся митозом. Задача этого периода увеличить число первичных половых клеток. Генетическая формула 2n2c.
3. Рост – сперматогонии и овогонии накапливают питательные вещества и увеличиваются в размерах. Теперь они называются сперматоциты I порядка и овоциты I порядка. В конце периода происходит репликация хромосом (2п2с ® 2n4с).
4. Созревание (мейоз) – происходит два последующих деления, между которыми нет интерфазы, и, следовательно, нет удвоения ДНК. Набор хромосом в клетках уменьшается в два раза, а набор хроматид уменьшается в 4 раза (мейоз I: 2n4с®1n2с, мейоз II: 2n2с®1n1с).
5. Формирование – клетки приобретают специфическое строение, обеспечивающее выполнение их функции. Этот период характерен только для сперматогенеза.
Отличия овогенеза от сперматогенеза
СПЕРМАТОГЕНЕЗ ОВОГЕНЕЗ
| 1. | Размножение сперматогоний начинается в эмбриональный период и продолжается до конца периода половой зрелости. | Размножение овогоний начинается и заканчивается в эмбриональный период. |
| 2. | Рост и созревание сперматозоидов происходит постоянно по достижении периода половой зрелости. | При овогенезе рост и первые стадии мейоза 1 происходят в эмбриогенезе. Мейоз 1 останавливается на стадии диплотены. Т.к. она растягивается во времени, её называют диктиотеной. С наступлением периода полового созревания, циклично 1 раз в месяц, 1 клетка заканчивает мейоз 1, и на стадии метафазы мейоза 2 происходит овуляция, т.е. выход яйцеклетки из яичника. Мейоз 2 заканчивается после оплодотворения. |
| 3. | В период роста сперматоциты 1 порядка меньше. | В период роста овоциты 1 порядка крупнее сперматоцитов. |
| 4. | Один сперматоцит 1 порядка дает начало четырём сперматозоидам одинакового размера. | Один овоцит 1 порядка дает начало одной крупной яйцеклетке и трём мелким полярным (направительным) тельцам, которые погибают. |
| 5. | Есть период формирования | Нет периода формирования |
4. Мейоз. Цитогенетическая характеристика. Биологическое значение мейоза.
Мейоз – это два последовательных деления клетки, приводящие к уменьшению числа хромосом вдвое. Каждое деление мейоза включает в себя 4 фазы: профаза, метафаза, анафаза и телофаза. Перед мейозом в конце периода роста происходит репликация ДНК, клетка имеет набор хромосом 2n4с.
Профаза I . Это самая продолжительная фаза мейоза I, во время которой происходят события, отличающие мейоз от митоза.
Профаза подразделяется на пять стадий, в неё вступают гаметоциты 1 порядка (2n4с)
1. лептотена – стадия тонких нитей. Начинается спирализация хромосом (хромосомы в световой микроскоп видны в виде нитей).
2. зиготена – стадия сливающихся нитей, гомологичные хромосомы отыскивают друг друга и объединяются. Этот процесс называется коньюгацией или синапсисом.
Механизм конъюгации:
а. в ДНК находятся многократно повторяющиеся последовательности, они обеспечивают точность прилегания гомологичных хромосом друг к другу по всей длине.
б. между гомологичными хромосомами образуется синаптонемальный комплекс из белков. Вдоль синаптонемального комплекса располагаются рекомбинационные узелки (в них находятся ферменты, которые участвуют в кроссинговере).
3. пахитена – стадия толстых нитей. Гомологичные хромосомы спирализованы и расположены близко друг к другу. Пару гомологичных хромосом называют – бивалент хромосом, или тетрада хроматид. Затем в определенных участках гомологичных хромосом происходит кроссинговер: перекрест гомологичных хромосом и обмен участками. При кроссинговере происходит разрыв двойной спирали ДНК, в одной отцовской хроматиде и одной материнской хроматиде, образовавшиеся участки соединяются наперекрест. Кроссинговер происходит в тех участках, где находятся рекомбинационные узелки синаптонемального комплекса. В конце пахитены синаптонемальный комплекс разрушается.
4. диплотена – стадия двойных нитей. Т. к. синаптонемальный комплекс разрушен, гомологичные хромосомы начинают отходить др. от др. Но они остаются связанными в точках кроссинговера. Эти участки хромосом называются хиазмы.
5. диакинез – хиазмы сдвигаются на концы хромосом, поэтому гомологичные хромосомы образуют кольцо.
Кроме того, в профазу 1 центриоли расходятся к разным полюсам клетки – образуется веретено деления. Разрушаются ядрышки, ядерная оболочка. В области центромеры с одной стороны каждой хромосомы образуются кинетохоры, от них отходят кинетохорные нити.
Метафаза 1. Конъюгированные хромосомы (биваленты) располагаются по экватору клетки, образуя метафазную пластинку. Заканчивается формирование веретена деления. В отличие от митоза нить веретена от каждой хромосомы направлена только к одному из полюсов. Это происходит потому, что из-за конъюгации каждая хромосома имеет только один кинетохор.
Анафаза 1. Гомологичные хромосомы, состоящие каждая из 2-х хроматид, отходят к противоположным полюсам. Расходятся хромосомы, а не хроматиды, т.к. центромера не делится, а хромосомная нить веретена тянется лишь к одному полюсу. Очень важно, что расхождение гомологичных хромосом происходит случайным образом. Поэтому к каждому полюсу отходит случайное число отцовских или материнских хромосом. На каждом полюсе находится в 2 раза меньше хромосом, чем было их в клетке до начала деления. Причем эти хромосомы качественно другие: большая часть каждой хромосомы – исходная хромосома; меньшая часть представлена заменённым участком гомологичной хромосомы. Так как гомологичные хромосомы – это отцовская и материнская хромосомы, можно сказать, что в результате кроссинговера образуются комбинированные хромосомы. В них содержатся новые комбинации отцовских и материнских генов, которые будут служить материалом для эволюционного процесса.
Tелофаза I – образуются 2 клетки (сперматоциты II-го порядка, овоциты II-го порядка) с набором 1n2с. Хромосомы деспирализуются, образуется ядерная оболочка, исчезает веретено деления, делится цитоплазма (цитокинез). Так как набор хромосом уменьшается вдвое, первое мейотическое деление называется редукционным.
После короткой интерфазы, во время которой не происходит репликации ДНК (т.к. отсутствует S-фаза), наступает мейоз II. Он называется эквационным, или уравнительным.
Мейоз II напоминает митоз, но особенность в том, что набор хромосом клеток, вступающий в мейоз II гаплоидный, и расходящиеся хроматиды содержат новую комбинацию генов по сравнению с хроматидами исходной клетки.
Итак, при сперматогенезе и овогенезе из одной исходной диплоидной клетки (2n4с) образуются четыре клетки с гаплоидным набором хромосом (1n1с), причем эти хромосомы содержат новую комбинацию генов.
Дата добавления: 2016-02-16; просмотров: 1955;