Механизмы окисления цитохрома в реакционном центре.
Поглощение кванта света в РЦ вызывает появление окисленного димера Р+ , который затем восстанавливается от донора (вторичного), роль которого в бактериальном фотосинтезе выполняют высоко- Сн и низкопотенциальные СL цитохромы.
Перенос электрона от цитохрома на Р+ явился важным источником информации о механизмах элементарных процессов в биологических структурах. Это дало
толчок новому направлению в биофизике фотосинтеза - исследованию транспорта электронов в биологических мембранах в условиях, исключающих обычную диффузию молекул переносчиков.
Окисление цитохромов в фотосинтетических мембранах не требует обычных активационных механизмов, подобных окислительно- восстановительным реакциям в растворах, а протекает по другим законам. Основополагающая концепция базируется на физическом явлении туннельного переноса электрона, который может происходить между молекулами донора Д и акцептора А в условиях, когда энергия электрона меньше высоты разделяющего их активационного барьера.
Основные положения теории туннельного переноса:
Согласно квантово-механическим представлениям существует определенная вероятность перекрывания электронных волновых функций начального и конечного состоянии, зависящая от высоты и ширины барьера. За время пребывания электрона на молекуле акцептора происходит потеря части электронной энергии и, как следствие, нарушение резонансного совпадения положения уровней А и Б. В результате этого обратное туннелирование практически невозможно, и перенос электрона становится необратимым. Диссипация электронной энергии за счет электроколебательных взаимодействий происходит за 10-12 -10-13 с с возбуждением колебательных акцептирующих мод в донорно-акцепторном комплексе в состоянии Б А*.
Таким образом, условием необратимости в конечном состоянии туннельного переноса является соотношение между временем электронного перехода тэл, зависящим от перекрывания волновых функций начального и конечного состояний, и временем колебательной релаксации т рел в конечном состоянии: тэл больше трел (золотое правило Ферми).
Типичные двухфазные температурные кривые окисления СL цитохрома, диаграмма потенциальных поверхностей электронной энергии Ui и Uf начального i и конечного f состояний донорно-акцепторной пары Д -А и экспериментальные данные о влиянии замещения H2O на D2O:
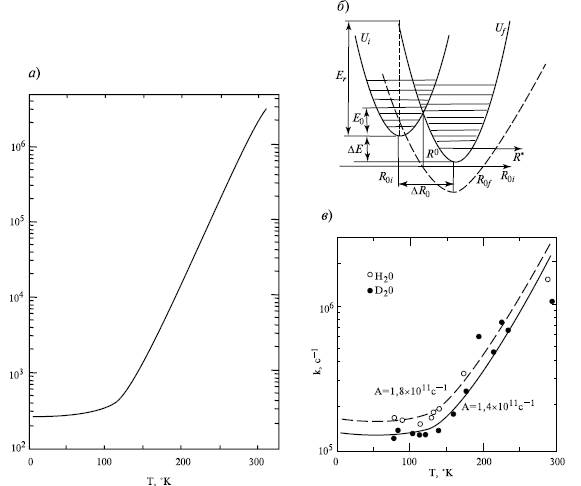
Температурная зависимость скорости окисления цитохрома в фотосинтезирующих бактериях (а); электронно-колебательные взаимодействия при туннелировании электрона (б); (в) —изотопный эффект замещения Н2О на Д2О; Ео —энергия активации, Еr—энергия реорганизации среды, дельта Е — тепловой эффект реакции переноса электрона, дельта Rо — смещение положения равновесия ядер при переходе из начального i в конечное состояние f. Штриховая кривая соответствует безактивационному переносу электрона Еа
Туннелирование электрона происходит, когда энергии состояний i и f становятся равными, что имеет место в точке R* ядерной координаты, где кривые Ui(R) и Uf(R) пересекаются: (Д*А)i->(Д*А)f
В состоянии (ДА*)f система оказывается на одном из высших колебательных подуровней, с которого происходит сброс избытка энергии с переводом на нижние колебательные подуровни и возбуждение акцептирующих мод. Приближение вдоль ядерной координаты к точке R* происходит при увеличении температуры и заселения высоких колебательных подуровней исходного состояния. При этом ширина активационного барьера перехода, как видно из риc б уменьшается, следовательно, скорость туннелирования растет с ростом температуры. При низких температурах в состоянии i заселены только нижние колебательные подуровни, и точка R* может быть достигнута только за счет туннелирования самих ядер (ядра — достаточно легкие частицы). Здесь, однако, ширина и высота барьера наибольшие, и поэтому скорость туннелирования минимальная, но от температуры уже не зависит. Таким образом, двухфазный характер температурной зависимости окисления цитохрома объясняется влиянием температуры на процесс перестройки ядерной конфигурации, который характеризуется величиной смещения дельта R = Rof - Roi исходной равновесной ядерной конфигурации Rof к новому равновесному значению Roi при переносе электрона. Разработанная количественная теория туннельного переноса в белковой среде дает выражения для константы скорости туннелирования в случае сильной и слабой электронно-колебательной связи, которые позволяют найти параметры процесса переноса электрона. Эти частоты характеризуются значениями ~ 300 - 400 с-1 . В качестве акцептирующей моды могут выступать колебания молекул воды в гидратных оболочках порфириновых молекул, в других случаях - также колебания атомов водорода в химических группах C—H, O—H, N—H.
Дата добавления: 2016-04-11; просмотров: 761;