Метаболизм углеводов
Конечными продуктами расщепления углеводов, поступающих в организм человека, являются глюкоза, фруктоза и галактоза, В печени фруктоза и галактоза превращаются в глюкозу, поэтому метаболизм глюкозы является общим конечным путем метаболизма всех углеводов.
Во всех клетках энергия запасается в виде АТФ, который образуется в ходе анаэробного (гликолиз) или аэробного (цикл лимонной кислоты) расщепления глюкозы. В печени и жировой ткани расщепление глюкозы может также происходить по пентозофосфатному пути, что позволяет не только получить энергию, но и выработать кофактор, играющий важную роль в синтезе жирных кислот. Глюкоза, поступающая в кровь после еды, запасается в организме в форме гликогена. Если депо гликогена насыщены, то поступающая глюкоза превращается в жир. Гликоген является легкодоступным источником глюкозы. Необходимость превращения глюкозы в гликоген при запасании энергетического материала обусловлена тем, что накопление легко растворимой глюкозы в клетках могло бы привести к осмотическому шоку с последующим разрушением клеточной мембраны. Нерастворимый гликоген осмотически неактивен. Только печень и, в меньшей степени, скелетные мышцы способны запасать значительное количество гликогена. Инсулин потенциирует синтез гликогена, тогда как адреналин и глюкагон, напротив, способствуют гликогенолизу. Поскольку в печени содержится около 70 г гликогена, а потребление глюкозы составляет в среднем 150 г/сут, запасы гликогена истощаются через 24 ч голодания. Чтобы обеспечить непрерывное снабжение органов и тканей глюкозой после 24 ч голодания, необходим синтез глюкозы de novo (глюконеогенез).
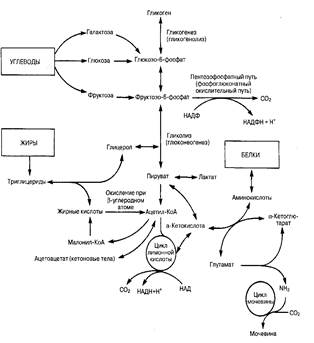
Рис. 34-3.Основные пути метаболизма в печени. Хотя небольшое количество АТФ образуется в результате промежуточных реакций, подавляющее большинство молекул АТФ синтезируется путем окислительного фосфорилирования восстановленных форм никотинамидаденинадинуклеотида (НАД) и никотинамидаденинадинуклеотидфосфата
Печень обладает уникальной способностью вырабатывать большое количество глюкозы из молочной и пировиноградной кислоты, аминокислот (в основном из аланина) и глицерола (образуемого в ходе метаболизма жиров). Нормальная концентрация глюкозы в крови поддерживается за счет глюконеогенеза в печени. Глюкокортикоиды, кате-холамины, глюкагон и тиреоидные гормоны потенциируют глюконеогенез, в то время как инсулин, напротив, ингибирует.
Метаболизм жиров
При насыщении депо углеводов избыток поступающих с пищей жиров (и белков) превращается в печени в жиры. Образующиеся жирные кислоты могут немедленно использоваться в качестве источника энергии или откладываться в жировой ткани или печени. В качестве источника энергии почти все клетки организма непосредственно утилизируют жирные кислоты, образующиеся из жиров пищи или синтезируемые в ходе промежуточного метаболизма углеводов и белков. Исключением являются эритроциты и мозговое вещество почки, где может утилизироваться только глюкоза. Нейроны в качестве источника энергии в обычных условиях используют только глюкозу, но через несколько дней голодания могут переключаться на жирные кислоты.
Жирные кислоты, образующиеся из жиров, вначале окисляются до ацетилкоэнзима А (ацетил-KoA), который, в свою очередь, окисляется в цикле лимонной кислоты с образованием АТФ. Печень обладает высокой способностью к окислению жирных кислот, в результате чего из избытка ацетил-KoA образуется ацетоацетат. Ацетоацетат, высвобождаемый гепатоцитами, служит альтернативным и легкодоступным (ацетоацетат быстро превращается в ацетил-КоА) циркулирующим в крови энергетическим субстратом для других видов клеток. Глюкагон усиливает окисление жирных кислот, а инсулин ингибирует его.
Ацетил-КоА, кроме того, используется печенью для образования холестерина и фосфолипидов, необходимых для синтеза клеточных мембран во всем организме. Синтезируемые в печени липопротеины имеют важное значение для транспорта липидов в крови.
Метаболизм белков
Печень играет ключевую роль в метаболизме белков. Если печень прекращает участвовать в метаболизме белков, то через несколько дней наступает смерть. Метаболизм белков осуществляется в несколько последовательных этапов: 1) дезаминиро-вание аминокислот; 2) образование мочевины (для элиминации аммиака, образующегося при дезами-нировании аминокислот); 3) взаимопревращения между заменимыми аминокислотами; 4) синтез белков плазмы.
Дезаминирование необходимо для превращения избытка аминокислот в углеводы и жиры. В ходе ферментативных процессов (чаще всего трансами-нирование) аминокислоты превращаются в соответствующие кетокислоты, а в качестве побочного продукта реакции образуется аммиак. Дезаминирование аланина весьма важно для глюконеогенеза в печени. Хотя дезаминирование может происходить в почках (в основном это касается глутамина, см. главу 30), основным местом дезаминирования в организме является печень. За исключением аминокислот, имеющих разветвленный радикал (лейцин, изолейцин и валин), в печени подвергаются де-заминированию почти все аминокислоты, поступающие в организм с белками пищи. Аминокислоты с разветвленным радикалом подвергаются метаболизму преимущественно в скелетных мышцах.
Аммиак, образующийся при дезаминировании аминокислот (а также вырабатывающийся под действием бактерий толстого кишечника и всасывающийся в кровь через стенку кишки), обладает высокой цитотоксичностью. В ходе нескольких последовательных реакций, проходящих в печени под действием ферментов, к двум молекулам аммиака присоединяется одна молекула СО2, в результате чего образуется мочевина. Образовавшаяся мочевина легко диффундирует из печени и затем выделяется через почки.
Трансаминирование соответствующих кетокис-лот в печени приводит к образованию заменимых аминокислот и компенсирует их недостаток в пищевом рационе. Незаменимые аминокислоты в соответствии со своим названием не синтезируются посредством этого механизма и должны поступать извне.
Почти все белки плазмы, за исключением имму-ноглобулинов, образуются в печени. В количественном отношении наиболее важными из этих белков являются альбумин и факторы свертывания. Альбумин обеспечивает поддержание нормального онкотического давления плазмы, а также является главным белком, осуществляющим связывание и транспорт гормонов и лекарственных препаратов. Следовательно, изменение концентрации альбумина оказывает влияние на концентрацию фармакологически активной, несвязанной фракции многих лекарственных препаратов.
Все факторы свертывания, за исключением фактора VIII и фактора фон Виллебранда, образуются в печени. Витамин К является необходимым кофактором для синтеза протромбина (фактор II) и факторов VІІ, IX и X. В печени синтезируется холинэ-стераза плазмы (синоним: псевдохолинэстераза) — фермент, который гидролизует эфиры, в том числе некоторые местные анестетики и сукцинилхолин. Другие важные белки, образующиеся в печени, включают ингибиторы протеаз (антитромбин III, А2-ан-типлазмин и А1-антитрипсин), транспортные белки (трансферрин, гаптоглобин и церулоплазмин), белки системы комплемента, А1-гликопротеин, С-реак-тивный белок и сывороточный амилоид типа А.
Дата добавления: 2016-03-30; просмотров: 1285;