Газообмен в легких и перенос газов кровью. 4 страница
Особое место в гуморальной регуляции деятельности дыхательного центра имеет изменение в крови напряжения СО2. При вдыхании газовой смеси, содержащей 5-7% СО2, увеличение парциального давления СО2 в альвеолярном воздухе задерживает выведение СО2 из
369


 венозной крови. Связанное с этим повышение напряжения СО2 в артериальной крови приводит к увеличению легочной вентиляции в 6-8 раз. Благодаря такому значительному увеличению объема дыхания, концентрация СО2 в альвеолярном воздухе возрастает не более, чем на 1%. Увеличение содержания СО2 в альвеолах на 0.2% вызывает увеличение вентиляции легких на 100%. Роль СО2 как главного регулятора дыхания, выявляется и в том, что недостаток содержания СО2 в крови понижает деятельность дыхательного центра и приводит к уменьшению объема дыхания и даже к полному прекращению дыхательных движения (апное). Это происходит, например, при искусственной гипервентиляции: произвольное увеличение глубины и частоты дыхания приводит к гипокапнии — снижению парциального давления СО2 в альвеолярном воздухе и артериальной крови. Поэтому после прекращения гипервентиляции появление очередного вдоха задерживается, а глубина и частота последующих вдохов вначале снижается.
венозной крови. Связанное с этим повышение напряжения СО2 в артериальной крови приводит к увеличению легочной вентиляции в 6-8 раз. Благодаря такому значительному увеличению объема дыхания, концентрация СО2 в альвеолярном воздухе возрастает не более, чем на 1%. Увеличение содержания СО2 в альвеолах на 0.2% вызывает увеличение вентиляции легких на 100%. Роль СО2 как главного регулятора дыхания, выявляется и в том, что недостаток содержания СО2 в крови понижает деятельность дыхательного центра и приводит к уменьшению объема дыхания и даже к полному прекращению дыхательных движения (апное). Это происходит, например, при искусственной гипервентиляции: произвольное увеличение глубины и частоты дыхания приводит к гипокапнии — снижению парциального давления СО2 в альвеолярном воздухе и артериальной крови. Поэтому после прекращения гипервентиляции появление очередного вдоха задерживается, а глубина и частота последующих вдохов вначале снижается.
Указанные изменения газового состава внутренней среды организма оказывают влияние на дыхательный центр опосредованно, через специальные хемочувствителъные рецепторы, расположенные непосредственно в структурах продолговатого мозга ("центральные хеморецепторы') и в сосудистых рефлексогенных зонах ("периферические хеморецепторы").
Центральными (медуллярными) хеморецепторами, постоянно участвующими в регуляции дыхания, называют нейрональные структуры в продолговатом мозге, чувствительные к напряжению СО2 и кислотно-щелочному состоянию омывающей их межклеточной мозговой жидкости. Хемочувствительные зоны имеются на переднебоковой поверхности продолговатого мозга около выходов подъязычного и блуждающего нервов в тонком слое мозгового вещества на глубине 0.2-0.4 мм. Медуллярные хеморецепторы постоянно стимулируются ионами водорода в межклеточной жидкости ствола мозга, концентрация которых зависит от напряжения СО2 в артериальной крови. Спинномозговая жидкость отделена от крови гемато-энцефалическим барьером, относительно непроницаемым для ионов Н+ и НСО3, но свободно пропускающим молекулярный СО2. При повышении напряжения СО2 в крови он диффундирует из кровеносных сосудов головного мозга в спинномозговую жидкость, в результате чего, в ней накапливаются ионы Н+, которые стимулируют медуллярные хеморецепторы. При повышении напряжения СО2 и концентрации водородных ионов в жидкости, омывающей медуллярные хеморецепторы, увеличивается активность инспираторных и падает активность экспираторных нейронов дыхательного центра продолговатого мозга. В результате этого, дыхание становится более глубоким и вентиляция легких растет, главным образом, за счет увеличения объема каждого вдоха. Напротив, снижение напряжения СО2 и подщелачивание межклеточной жидкости ведет к полному или частичному исчезновению реакции увеличения объема дыхания на избыток СО2 (гиперкапнию) и ацидоз, а также к резкому угнетению инспираторной активности дыхательного центра вплоть до остановки дыхания.
370


 Периферические хеморецепторы, воспринимающие газовый состав артериальной крови, расположены в двух областях: дуге аорты и месте деления (бифуркация) общей сонной артерии (каротидный синус), т.е. в тех же зонах, что и барорецепторы, реагирующие на изменения кровяного давления. Однако, хеморецепторы представляют собой самостоятельные образования, заключенные в особых тельцах — клубочках или гломусах, которые находятся вне сосуда. Афферентные волокна от хеморецепторов идут: от дуги аорты — в составе аортальной ветви блуждающего нерва, а от синуса сонной артерии — в каротидной ветви языкоглоточного нерва, так называемом нерве Геринга. Первичные афференты синусного и аортального нерва проходят через ипсилатеральное ядро солитарного тракта. Отсюда хеморецептивные импульсы поступают к дорсальной группе дыхательных нейронов продолговатого мозга.
Периферические хеморецепторы, воспринимающие газовый состав артериальной крови, расположены в двух областях: дуге аорты и месте деления (бифуркация) общей сонной артерии (каротидный синус), т.е. в тех же зонах, что и барорецепторы, реагирующие на изменения кровяного давления. Однако, хеморецепторы представляют собой самостоятельные образования, заключенные в особых тельцах — клубочках или гломусах, которые находятся вне сосуда. Афферентные волокна от хеморецепторов идут: от дуги аорты — в составе аортальной ветви блуждающего нерва, а от синуса сонной артерии — в каротидной ветви языкоглоточного нерва, так называемом нерве Геринга. Первичные афференты синусного и аортального нерва проходят через ипсилатеральное ядро солитарного тракта. Отсюда хеморецептивные импульсы поступают к дорсальной группе дыхательных нейронов продолговатого мозга.
Артериальные хеморецепторы вызывают рефлекторное увеличение легочной вентиляции в ответ на снижение напряжения кислорода в крови (гипоксемию). Даже в обычных {нормоксических) условиях эти рецепторы находятся в состоянии постоянного возбуждения, которое исчезает только при вдыхании человеком чистого кислорода. Уменьшение напряжения кислорода в артериальной крови ниже нормального уровня вызывает усиление афферентации из аортальных и синокаротидных хеморецепторов. Вдыхание гипоксической смеси ведет к учащению и увеличению регулярности импульсов, посылаемых хеморецепторами каротидного тельца.
Повышению напряжения СО2 артериальной крови и соответствующему подъему вентиляции также сопутствует рост импульсной активности, направляемой в дыхательный центр от хеморецепторов каротидного синуса. Особенность роли, которую играют артериальные хеморецепторы в контроле за напряжением углекислоты, состоит в том, что они ответственны за начальную, быструю, фазу вентиляторной реакции на гиперкапнию. При их денервации указанная реакция наступает позднее и оказывается более вялой, так как развивается в этих условиях лишь после того, как повысится напряжение СО2 области хемочувствительных мозговых структур.
Гиперкапническая стимуляция артериальных хеморецепторов, подобно гипоксической, носит постоянный характер. Эта стимуляция начинается при пороговом напряжении СО2 20-30 мм рт.ст и, следовательно, имеет место уже в условиях нормального напряжения СО2 в артериальной крови (около 40 мм рт.ст.).
Важным моментом для регуляции дыхания является взаимодействие гуморальных стимулов дыхания. Оно проявляется, например, в том, что на фоне повышенного артериального напряжения СО2 или увеличенной концентрации водородных ионов вентиляторная реакция на гипоксемию становится интенсивнее. Поэтому снижение парциального давления кислорода и одновременное повышение парциального давления углекислого газа в альвеолярном воздухе вызывают нарастание легочной вентиляции, превышающее арифметическую сумму ответов, которые вызывают эти факторы, действуя порознь. Физиологическое значение этого явления заключается в том,
371




 что указанное сочетание стимуляторов дыхания имеет место при мышечной деятельности, которая сопряжена с максимальным подъемом газообмена и требует адекватного ему усиления работы дыхательного аппарата.
что указанное сочетание стимуляторов дыхания имеет место при мышечной деятельности, которая сопряжена с максимальным подъемом газообмена и требует адекватного ему усиления работы дыхательного аппарата.
Установлено, что гипоксемия снижает порог и увеличивает интенсивность вентиляторной реакции на СО2. Однако, у человека при недостатке кислорода во вдыхаемом воздухе увеличение вентиляции происходит лишь при условии, когда артериальное напряжение СО2 составляет не менее 30 мм рт.ст. При уменьшении парциального давления О2 во вдыхаемом воздухе (например, при дыхании газовыми смесями с низким содержанием О2, при пониженном атмосферном давлении в барокамере или в горах) возникает гипервентиляция, направленная на предупреждение значительного снижения парциального давления О2 в альвеолах и напряжения его в артеальной крови. При этом из-за гипервентиляции наступает снижение парциального давления СО2 в альвеолярном воздухе и развивается ги-покапния, приводящая к уменьшению возбудимости дыхательного центра. Поэтому при гипоксической гипоксии, когда парциальное давление СО2 во вдыхаемом воздухе снижается до 12 кПа (90 мм рт.ст.) и ниже, система регуляции дыхания может лишь частично обеспечить поддержание напряжения О2 и СО2 на должном уровне. В этих условиях, несмотря на гипервентиляцию, напряжение О2 все же снижается, и возникает умеренная гипоксемия.
В регуляции дыхания функции центральных и периферических рецепторов постоянно дополняют друг друга и, в общем, проявляют синергизм. Так, импульсация хеморецепторов каротидного тельца усиливает эффект стимуляции медуллярных хемочувствительных структур. Взаимодействие центральных и периферических хеморецепторов имеет жизненно важное значение для организма, например, в условиях дефицита О2. При гипоксии из-за снижения окислительного метаболизма в мозге чувствительность медуллярных хеморецепторов ослабевает или исчезает, вследствие чего снижается активность дыхательных нейронов. Дыхательный центр в этих условиях получает интенсивную стимуляцию от артериальных хеморецепторов, для которых гипоксемия является адекватным раздражителем. Таким образом, артериальные хеморецепторы служат "аварийным" механизмом реакции дыхания на изменение газового состава крови, и, прежде всего, на дефицит кислородного снабжения мозга.
Взаимосвязь регуляции внешнего дыхания и других функций организма.Обмен газов в легких и тканях и приспособление его к запросам тканевого дыхания при различных состояниях организма обеспечивается путем изменения не только легочной вентиляции, но и кровотока как в самих легких, так и других органах. Поэтому механизмы нейрогуморальной регуляции дыхания и кровообращения осуществляются в тесном взаимодействии. Рефлекторные влияния, исходящие из рецептивных полей сердечно-сосудистой системы (например, гинокаротидной зоны), изменяют деятельность как дыха-

 тельного, так и сосудодвигательного центров. Нейроны дыхательного центра подвержены рефлекторным воздействиям со стороны бароре-цепторных зон сосудов — дуги аорты, каротидного синуса. Сосудо-двигательные рефлексы неразрывно связаны и с изменением функции дыхания. Повышение сосудистого тонуса и усиление сердечной деятельности, соответственно, сопровождаются усилением дыхательной функции. Например, при физической или эмоциональной нагрузке у человека обычно имеет место согласованное повышение минутного объема крови в большом и малом круге, артериального давления и легочной вентиляции. Однако, резкое повышение артериального давления вызывает возбуждение синокаротидных и аортальных барорецепторов, которое приводит к рефлекторному торможению дыхания. Понижение артериального давления, например, при кровопотере, приводит к увеличению легочной вентиляции, что вызвано, с одной стороны, снижением активности сосудистых барорецепторов, с другой — возбуждением артериальных хеморецепторов в результате местной гипоксии, вызванной уменьшением в них кровотока. Учашение дыхания возникает пи повышении давления крови в малом круге кровообращения и при растяжении левого предсердия.
тельного, так и сосудодвигательного центров. Нейроны дыхательного центра подвержены рефлекторным воздействиям со стороны бароре-цепторных зон сосудов — дуги аорты, каротидного синуса. Сосудо-двигательные рефлексы неразрывно связаны и с изменением функции дыхания. Повышение сосудистого тонуса и усиление сердечной деятельности, соответственно, сопровождаются усилением дыхательной функции. Например, при физической или эмоциональной нагрузке у человека обычно имеет место согласованное повышение минутного объема крови в большом и малом круге, артериального давления и легочной вентиляции. Однако, резкое повышение артериального давления вызывает возбуждение синокаротидных и аортальных барорецепторов, которое приводит к рефлекторному торможению дыхания. Понижение артериального давления, например, при кровопотере, приводит к увеличению легочной вентиляции, что вызвано, с одной стороны, снижением активности сосудистых барорецепторов, с другой — возбуждением артериальных хеморецепторов в результате местной гипоксии, вызванной уменьшением в них кровотока. Учашение дыхания возникает пи повышении давления крови в малом круге кровообращения и при растяжении левого предсердия.
На работу дыхательного центра оказывает влияние афферентация от периферических и центральных терморецепторов, особенно при резких и внезапных температурных воздействиях на рецепторы кожи. Погружение человека в холодную воду, например, тормозит выдох, в результате чего возникает затяжной вдох. У животных, у которых отсутствуют потовые железы (например, у собаки), с повышением температуры внешней среды и ухудшением теплоотдачи увеличивается вентиляция легких за счет учашения дыхания (температурное полипное) и усиливается испарение воды через систему дыхания.
Рефлекторные влияния на дыхательный центр весьма обширны, и практически все рецепторные зоны при их раздражении изменяют дыхание. Эта особенность рефлекторной регуляции дыхания отражает общий принцип нейронной организации ретикулярной формации ствола мозга, в состав которой входит и дыхательный центр. Нейроны ретикулярной формации, в том числе и дыхательные нейроны, имеют обильные коллатерали почти от всех афферентных систем организма, что и обеспечивает, в частности, разносторонние рефлекторные влияния на дыхательный центр. На деятельности нейронов дыхательного центра отражается большое количество различных неспецифических рефлекторных влияний. Так, болевые раздражения сопровождаются немедленным изменением дыхательной ритмики. Функция дыхания теснейшим образом связана с эмоциональными процессами: почти все эмоциональные проявления человека сопровождаются изменением функции дыхания; смех, плач — это измененные дыхательные движения.
В дыхательный центр продолговатого мозга непосредственно поступает импульсация от рецепторов легких и рецепторов крупных сосудов, т.е. рецептивных зон, раздражение которых имеет особенно существенное значение для регуляции внешнего дыхания. Однако,
373



 для адекватного приспособления функции дыхания к меняющимся условиям существования организма система регуляции должна обладать полной информацией о том, что происходит в организме и в окружающей среде. Поэтому для регуляции дыхания имеют значение все афферентные сигналы от разнообразных рецептивных полей организма. Однако, вся эта сигнализация поступает не непосредственно в дыхательный центр продолговатого мозга, а в различные уровни головного мозга (рис.8.10), и от них непосредственно может передаваться как на дыхательную, так и на другие функциональные системы. Различные центры головного мозга образуют с дыхательным центром функционально подвижные ассоциации, обеспечивающие полноценное регулирование дыхательной функции.
для адекватного приспособления функции дыхания к меняющимся условиям существования организма система регуляции должна обладать полной информацией о том, что происходит в организме и в окружающей среде. Поэтому для регуляции дыхания имеют значение все афферентные сигналы от разнообразных рецептивных полей организма. Однако, вся эта сигнализация поступает не непосредственно в дыхательный центр продолговатого мозга, а в различные уровни головного мозга (рис.8.10), и от них непосредственно может передаваться как на дыхательную, так и на другие функциональные системы. Различные центры головного мозга образуют с дыхательным центром функционально подвижные ассоциации, обеспечивающие полноценное регулирование дыхательной функции.
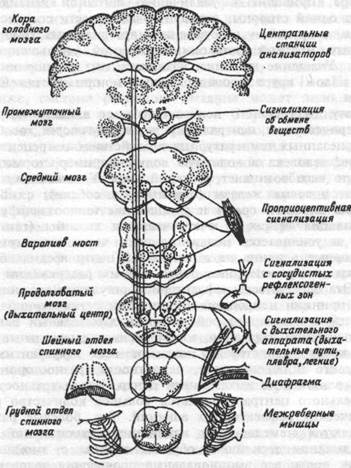
Рис.8.10. Схема организации центрального аппарата регуляции дыхания.
Стрелками обозначены пути передачи регулирующих влияний к дыхательному центру продолговатого мозга.

 Как видно на рис. 8.10, в центральный механизм, регулирующий дыхание, включены разные уровни ЦНС. Значение для регуляции дыхания структур стволовой части мозга, в том числе варолиевого моста, среднего мозга, заключается в том, что эти отделы ЦНС получают и переключают на дыхательный центр проприоцептивную и интероцептивную сигнализацию, а промежуточный мозг — сигнализацию об обмене веществ. Кора больших полушарий, как центральная станция анализаторных систем, вбирает и обрабатывает сигналы от всех органов и систем, делая возможным адекватное приспособление различных функциональных систем, в том числе и дыхания, к тончайшим изменениям жизнедеятельности организма.
Как видно на рис. 8.10, в центральный механизм, регулирующий дыхание, включены разные уровни ЦНС. Значение для регуляции дыхания структур стволовой части мозга, в том числе варолиевого моста, среднего мозга, заключается в том, что эти отделы ЦНС получают и переключают на дыхательный центр проприоцептивную и интероцептивную сигнализацию, а промежуточный мозг — сигнализацию об обмене веществ. Кора больших полушарий, как центральная станция анализаторных систем, вбирает и обрабатывает сигналы от всех органов и систем, делая возможным адекватное приспособление различных функциональных систем, в том числе и дыхания, к тончайшим изменениям жизнедеятельности организма.
Своеобразие функции внешнего дыхания заключается в том, что она в одной и той же мере и автоматическая, и произвольно управляемая. Человек прекрасно дышит во сне и под наркозом; у животных дыхание сохраняет практически нормальный характер даже после удаления всего переднего мозга. В то же время любой человек может произвольно, хотя и ненадолго, остановить дыхание или изменить его глубину и частоту. Произвольное управление дыханием основано на наличии в коре больших полушарий представительства дыхательных мышц и наличии корковомедуллярных нисходящих активирующих и тормозных влияний на эфферентную часть дыхательного центра. Возможность произвольного управления дыханием ограничена определенными пределами изменений напряжения кислорода и углекислоты, а также рН крови. При чрезмерной произвольной задержке дыхания или резком отклонении фактического минутного объема вентиляции от физиологически обоснованного возникает стимул, который возвращает дыхание под контроль дыхательного центра, преодолевая корковое влияние.
Роль коры головного мозга в регуляции дыхания показана в экспериментах на животных с электрическим раздражением различных зон больших полушарий, а также с их удалением. Оказалось, что стоит лишь бескорковому животному в течение 1-2 мин сделать несколько шагов, как у него начинается резко выраженная и длительная одышка, т.е. значительное учащение и усиление дыхания. Следовательно, если требуется приспособление дыхания к условиям внешней среды, например при мышечной деятельности, необходимо участие высших отделов центральной нервной системы. Бескорковые животные сохраняют равномерное дыхание лишь в состоянии полного покоя и теряют способность к адаптации дыхания к изменениям внешней среды при мышечной работе.
Влияние коры головного мозга на дыхание у человека проявляется, например, в усилении дыхания еще в стартовых условиях перед выполнением мышечных усилий, сразу после команды "приготовиться". Дыхание усиливается у человека непосредственно после начала движений, когда образующиеся при мышечной работе гуморальные вещества еще не достигли дыхательного центра. Следовательно, усиление дыхания в самом начале мышечной работы обусловлено рефлекторными воздействиями, повышающими возбудимость дыхательного центра.
375


 Кортикальные влияния на дыхание отчетливо проявляются при тренировке к выполнению одной и той же работы: при этом происходит постепенное развитие и совершенствование адекватных для данной работы функциональных взаимосвязей между мышечной работой и дыханием. На это указывает динамика изменения внешнего дыхания в процессе, например, тренировки к работе на велоэрго-метре с переменной интенсивностью. Если темп работы постоянен, а ее интенсивность периодически меняется по заранее составленному графику, то по мере тренировки с такой программой средний уровень легочной вентиляции снижается, но изменение вентиляции при переключении на новый уровень интенсивности наступает быстрее. Следовательно, в результате тренировки к работе переменной интенсивности развивается способность к более быстрому переключению деятельности дыхательного аппарата на новый уровень функциональной активности, адекватной новым условиям работы. Лучшая согласованность во времени процессов координации функции внешнего дыхания при переходе от одних условий работы к другим связана с функциональной перестройкой высших отделов ЦНС. В результате этого, по мере тренировки к мышечной работе колебания объема дыхания становятся меньше и дыхание делается более ровным. Выработанный, таким образом, динамический стереотип проявляется в том, что при переходе к работе с постоянной интенсивностью вентиляция легких имеет выраженный волнообразный характер.
Кортикальные влияния на дыхание отчетливо проявляются при тренировке к выполнению одной и той же работы: при этом происходит постепенное развитие и совершенствование адекватных для данной работы функциональных взаимосвязей между мышечной работой и дыханием. На это указывает динамика изменения внешнего дыхания в процессе, например, тренировки к работе на велоэрго-метре с переменной интенсивностью. Если темп работы постоянен, а ее интенсивность периодически меняется по заранее составленному графику, то по мере тренировки с такой программой средний уровень легочной вентиляции снижается, но изменение вентиляции при переключении на новый уровень интенсивности наступает быстрее. Следовательно, в результате тренировки к работе переменной интенсивности развивается способность к более быстрому переключению деятельности дыхательного аппарата на новый уровень функциональной активности, адекватной новым условиям работы. Лучшая согласованность во времени процессов координации функции внешнего дыхания при переходе от одних условий работы к другим связана с функциональной перестройкой высших отделов ЦНС. В результате этого, по мере тренировки к мышечной работе колебания объема дыхания становятся меньше и дыхание делается более ровным. Выработанный, таким образом, динамический стереотип проявляется в том, что при переходе к работе с постоянной интенсивностью вентиляция легких имеет выраженный волнообразный характер.
Роль высших отделов ЦНС в регуляции дыхания у человека проявляется не только в его способности произвольно менять темп, ритм и амплитуду дыхательных движения, но и в его способности к "сознательному" восприятию своего гипоксического, либо гипер-капнического состояния.
Человек не может непосредственно воспринимать содержание кислорода и углекислого газа во вдыхаемом воздухе в силу отсутствия адекватных рецепторов в дыхательных путях и легких. Однако, с помощью метода активного выбора предпочитаемых дыхательных смесей (так называемый газопреферендум) показано, что люди избегают дышать газовыми смесями, которые вызывают в организме гипоксические или гиперкапнические сдвиги. Например, человеку предлагали выбрать одну из двух поочередно вдыхаемых смесей газов с разным, неизвестным ему содержанием кислорода. В таких условиях смеси, содержавшие 15% О2 и более, люди еще не отличали от обычного воздуха, 12%-е содержание кислорода вызывало у части людей уже отрицательную реакцию, а смесь с 9% кислорода отвергалась почти всеми испытуемыми. Аналогичным образом человек, избегал дышать смесями, обогащенными углекислым газом.
Исследования на спортсменах выявили их способность оценивать гипоксические и гиперкапнические сдвиги в своем организме не только при вдыхании соответствующих газов, но и при интенсивной мышечной деятельности. В частности, после спортивной тренировки исследуемые могли по своим ощущениям почти точно определять степень оксигенации собственной артериальной крови.
376

 При дыхании газовыми смесями, имеющими физиологически неадекватный состав, человек независимо от интенсивности развивающейся гипервентиляции иногда заявляет, что ему "трудно дышать", т.е. жалуется на одышку. Ощущение одышки является отражением рассогласования между хеморецептивной сигнализацией и другими звеньями рефлекторной регуляции дыхания, в том числе обратной афферентацией, исходящей из работающей дыхательной мускулатуры. Такого рода ощущения лежат в основе самоконтроля резервной работоспособности при выполнении человеком значительной мышечной нагрузки.
При дыхании газовыми смесями, имеющими физиологически неадекватный состав, человек независимо от интенсивности развивающейся гипервентиляции иногда заявляет, что ему "трудно дышать", т.е. жалуется на одышку. Ощущение одышки является отражением рассогласования между хеморецептивной сигнализацией и другими звеньями рефлекторной регуляции дыхания, в том числе обратной афферентацией, исходящей из работающей дыхательной мускулатуры. Такого рода ощущения лежат в основе самоконтроля резервной работоспособности при выполнении человеком значительной мышечной нагрузки.
Дата добавления: 2016-03-27; просмотров: 1585;