Газообмен в легких и перенос газов кровью. 1 страница
Функции дыхания
Общим для всех живых клеток является процесс расщепления органических молекул последовательным рядом ферментативных реакций, в результате чего высвобождается энергия. Практически любой процесс, при котором окисление органических веществ ведет к. выделению химической энергии, называют дыханием. Если для него требуется кислород, то дыхание называют аэробным, а если же реакции идут в отсутствии кислорода — анаэробным дыханием. Для всех тканей позвоночных животных и человека основным источником энергии являются процессы аэробного окисления, которые протекают в митохондриях клеток, приспособленных для превращения энергии окисления в энергию резервных макроэргических соединений типа АТФ. Последовательность реакций, посредством которых клетки организма человека используют энергию связей органических молекул, называется внутренним, тканевым или клеточным дыханием.
Под дыханием высших животных и человека понимают совокупность процессов, обеспечивающих поступление во внутреннюю среду организма кислорода, использование его для окисления органических веществ и удаление из организма углекислого газа.
Функцию дыхания у человека реализуют: 1) внешнее, или легочное, дыхание, осуществляющее газообмен между наружной и внутренней средой организма (между воздухом и кровью); 2) кровообращение, обеспечивающее транспорт газов к тканям и от них; 3) кровь как специфическая газотранспортная среда; 4) внутреннее, или тканевое, дыхание, осуществляющее непосредственный процесс клеточного окисления; 5) средства нейрогуморальной регуляции дыхания.
Результатом деятельности системы внешнего дыхания является обогащение крови кислородом и освобождение от избытка углекислоты. Изменение газового состава крови в легких обеспечивают три процесса; 1) непрерывная вентиляция альвеол для поддержания нормального газового состава альвеолярного воздуха; 2) диффузия газов через альвеолярно- капиллярную мембрану в объеме, достаточном для достижения равновесия давления кислорода и углекислого газа в альвеолярном воздухе и крови; 3) непрерывный кровоток в капиллярах легких в соответствии с объемом их вентиляции
* В подготовке главы принимал участие Л.В.Надежкин.
340
8.1. Дыхательный акт и вентиляция легких.
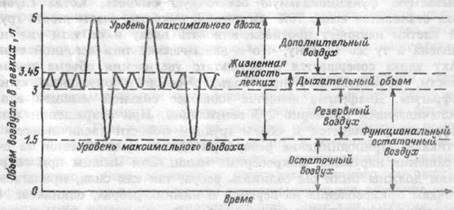
Количество воздуха, находящееся в легких после максимального вдоха, составляет общую емкость легких, величина которой у взрослого человека составляет 4200-6000 мл (рис.8.1). Она состоит из жизненной емкости легких, представляющей собой то количество воздуха (3300-4800 мл), которое выходит из легких при максимально глубоком выдохе после максимально глубокого вдоха, и остаточного воздуха (1100-1200 мл), который еще остается в легких после максимального выдоха
Жизненная емкость составляет три легочных объема: дыхательный объем, представляющий собой объем (400- 500 мл) воздуха, вдыхаемый и выдыхаемый при каждом дыхательном цикле; резервный объем вдоха (дополнительный воздух), т.е. тот объем (1900-3300 мл) воздуха, который можно вдохнуть при максимальном вдохе после обычного вдоха; резервный объем выдоха (резервный воздух), т.е. объем (700- 1000 мл), который можно выдохнуть при максимальном выдохе после обычного выдоха. При спокойном дыхании после выдоха в легких остается резервный объем выдоха и остаточный объем. Сумму этих объемов называют функциональной остаточной емкостью, а также нормальной емкостью легких, емкостью покоя, емкостью равновесия, буферным воздухом.
| Рис.8.1. Легочные объемы и емкости. |
Биомеханика дыхательного акта.Аппарат вентиляции состоит из двух анатомо-физиологических образований: грудной клетки с дыхательными мышцами и легких с дыхательными путями. Грудной отдел позвоночника и грудины с укрепленными на них 12 парами ребер и дыхательными мышцами наряду с диафрагмой образуют жесткий, подвижный, обладающий эластичностью футляр для легких, который изменяет свой объем вследствие сокращений дыхательных мышц.
341




 Дыхательные мышцы, благодаря которым осуществляются периодические изменения объема грудной клетки, относятся к поперечнополосатой скелетной мускулатуре, но они отличаются от других скелетных мышц. Во-первых, это единственные скелетные мышцы, от которых зависит жизнь; поэтому на протяжении всей жизни они должны ритмически сокращаться. Во-вторых, они находятся как под произвольным, так и непроизвольным контролем.
Дыхательные мышцы, благодаря которым осуществляются периодические изменения объема грудной клетки, относятся к поперечнополосатой скелетной мускулатуре, но они отличаются от других скелетных мышц. Во-первых, это единственные скелетные мышцы, от которых зависит жизнь; поэтому на протяжении всей жизни они должны ритмически сокращаться. Во-вторых, они находятся как под произвольным, так и непроизвольным контролем.
Различают основные и вспомогательные дыхательные мышцы. К первым относят диафрагму и межреберные мышцы, обеспечивающие вентиляцию легких в физиологических условиях. К вспомогательным относят мышцы шеи, часть мышц верхнего плечевого пояса, мышцы брюшного пресса, принимающие участие в форсированном вдохе или выдохе в обстоятельствах, затрудняющих вентиляцию легких. Легкие, находящиеся внутри грудной клетки, отделены от ее стенок плевральной полостью (щелью). В грудной клетке они находятся в растянутом состоянии. За счет того, что легкие обладают эластичностью (эластичность — сочетание растяжимости и упругости), давление в межплевральном щелевидном пространстве (так называемое плевральное давление) меньше альвеолярного на величину, обусловленную эластической тягой легких. Давление в плевральной щели может быть измерено путем прокола грудной стенки полой иглой, соединенной с манометром. Как только игла попадает в плевральное пространство, манометр показывает давление ниже атмосферного. Плевральное давление поэтому часто называют отрицательным, принимая уровень атмосферного давления за нуль V. После спокойного выдоха оно ниже атмосферного примерно на 6 мм рт.ст., а во время спокойного вдоха — на 9 мм рт.ст.
Отрицательное давление в плевральной полости стремится сжать грудную клетку, а эластическая тяга самой грудной клетки направлена в противоположную от тяги легких сторону (что облегчает вдох). Соотношение указанных сил определяет уровень спокойного дыхания и величину объема воздуха в легких после выдоха — так называемую функциональную остаточную емкость. Когда глубина вдоха становится выше 70% жизненной емкости, эластичность грудной клетки начинает противодействовать вдоху и ее тяга уже направлена в ту же сторону, что и эластическая тяга легочной ткани.
Акт вдоха совершается в результате увеличения объема грудной полости, происходящего при подъеме ребер и опускании купола диафрагмы. Диафрагма является наиболее сильной мышцей вдоха, обеспечивающей примерно 2/3 вентиляции. При сокращении купол диафрагмы уплощается и объем грудной полости увеличивается по вертикали. Приподнимание ребер при вдохе осуществляется за счет сокращения наружных межреберных мышц. Эти мышцы при сокращении должны были бы сближать ребра, так как сила, прилагаемая к точкам прикрепления на верхних и нижних ребрах, одинакова. Но при косом расположении этих мышц плечо и момент силы у верхнего ребра всегда меньше, чем у нижнего. Поэтому при сокращении мышц ребра приподнимаются, что увеличивает сечение грудной клетки как в передне-заднем, так и ь поперечном направлении.
342


 Увеличение объема грудной клетки при сокращении мышц вдоха приводит к уменьшению давления в плевральной полости. В результате этого воздух в легких расширяется, а давление его становится ниже атмосферного. Вследствие образующейся разности между давлением в окружающей среде и в альвеолах наружный воздух поступает по трахеобронхиальным путям в альвеолы.
Увеличение объема грудной клетки при сокращении мышц вдоха приводит к уменьшению давления в плевральной полости. В результате этого воздух в легких расширяется, а давление его становится ниже атмосферного. Вследствие образующейся разности между давлением в окружающей среде и в альвеолах наружный воздух поступает по трахеобронхиальным путям в альвеолы.
Во время вдоха мышцы преодолевают ряд сил: 1) эластическое сопротивление грудной клетки (после достижения 70% жизненной емкости) и внутренних органов, отдавливаемых книзу диафрагмой; 2) эластическое сопротивление легких; 3) динамическое (вязкое) сопротивление всех перемещаемых тканей; 4) аэродинамическое сопротивление дыхательных путей; 5) тяжесть перемещаемой части грудной клетки; 6) силы, обусловленные инерцией перемещаемых масс.
Энергия мышц, затраченная на преодоление всех видов динамического сопротивления (обусловленного трением), переходит в тепло и в дальнейшем процессе дыхания не участвует. Остальная часть энергии мышц переходит в потенциальную энергию растяжения всех эластических тканей и потенциальную энергию тяжести перемещаемой части грудной клетки. При расслаблении мышц вдоха под действием эластических сил грудной клетки и внутренних органов и силы тяжести грудной клетки ее объем уменьшается — происходит выдох, который при спокойном дыхании является пассивным актом. При активном форсированном выдохе к перечисленным силам присоединяется сокращение внутренних межреберных мышц и мышц брюшного пресса.
Уменьшение объема грудной клетки при выдохе приводит к повышению плеврального давления. В результате этого и под действием эластической тяги легких воздух в альвеолах сжимается, его давление становится выше атмосферного, и он начинает выходить наружу. Когда эластическая тяга легких уравновесится понижающимся давлением в плевральной полости, выдох заканчивается. Таким образом, действие дыхательных мышц на легкие осуществляется не непосредственно, а через изменение давления в плевральной полости. Непосредственной же причиной движения воздуха через дыхательные пути при вдохе и выдохе являются колебания альвеолярного давления.
В зависимости от того, связано ли расширение грудной клетки при нормальном дыхании преимущественно с поднятием ребер или уплощением диафрагмы, различают грудной (реберный) и брюшной типы дыхания. При грудном типе дыхание обеспечивается, в основном, за счет работы межреберных мышц, а диафрагма смещается пассивно в соответствии с изменением внутригрудного давления. При брюшном типе дыхания в результате мощного сокращения диафрагмы не только понижается внутриплевральное давление, но и одновременно повышается внутрибрюшное. Этот тип дыхания более эффективен, так как при нем сильнее вентилируются легкие и облегчается венозный возврат крови от органов брюшной полости к сердцу.
343




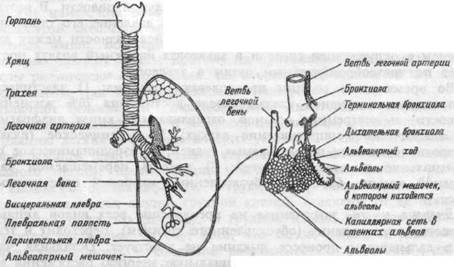
Рис.8.2. Схема строения легких человека.
 Если вскрыть грудную клетку — произвести пневмоторакс, то стремление легких к спадению за счет их эластической тяги проявится в полной мере — легкие займут так называемый объем коллапса, который значительной меньше, чем остаточный объем. Вентиляция легких при разгерметизации плевральной полости становится невозможной, поскольку при изменении объема грудной клетки воздух движется не через дыхательные пути, а через искусственно созданное отверстие в стенке грудной клетки.
Если вскрыть грудную клетку — произвести пневмоторакс, то стремление легких к спадению за счет их эластической тяги проявится в полной мере — легкие займут так называемый объем коллапса, который значительной меньше, чем остаточный объем. Вентиляция легких при разгерметизации плевральной полости становится невозможной, поскольку при изменении объема грудной клетки воздух движется не через дыхательные пути, а через искусственно созданное отверстие в стенке грудной клетки.
Соответственно, структурным и функциональным характеристикам легкие делят на воздухопроводящие (дыхательные) пути и альвеолы, составляющие респираторную зону, в которой непосредственно осуществляется газообмен (рис.8.2).
Основная функция воздухопроводящих путей состоит в доставке воздуха в респираторную зону. Воздухоносные пути подразделяют на верхние и нижние. К верхним относят носовые ходы, полость рта, носоглотку, придаточные пазухи носа, к нижним — гортань, трахею и все бронхи, вплоть до их конечных ответвлений. Узким местом в гортани является голосовая щель, имеющая максимальную ширину около 7 мм. При вдохе голосовая щель расширяется, а при выдохе — сужается.
Трахея у взрослого человека имеет длину около 12 см и диаметр 16-27 мм. На уровне 5-го грудного позвонка она делится на правый и левый главные бронхи, которые затем последовательно делятся по типу дихотомии. Каждое последовательное деление ветвей бронхи-альнго дерева по типу дихотомии образует как бы новое поколение (генерацию) элементов дыхательного тракта. Всего у человека от трахеи до альвеол имеется примерно 23 таких генерации (рис.8.3).
344
 | |||||||
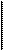 | |||||||
 | |||||||
 | |||||||
|
Трахея и следующие за ней 16 генерации бронхов и бронхиол относят к проводящей (кондуктивкой) зоне легких, т.е. зоне, в которой отсутствует контакт между воздухом и легочным капилля-рами и которую называют поэтому анатомическим мертвым пространством. Объем этой зоны составляет около 175 мл. Суммарная площадь поперечного сечения всех бронхиол 16-й генерации равно 180 см2, т.е. превышает плошадь поперечного сечения трахеи (2.54 см2) более, чем в 70 раз.
| Рис.8.3. Схема воздухоносных путей легких человека. |
Последующие три (17,18,19) генерации бронхиол (дыхательные бронхиолы) относят к переходной (транзиторной) зоне, где наряду с проведением воздуха начинает осуществляться и газообмен в малочисленных альвеолах, расположенных на поверхности дыхательных бронхиол. Их количество составляет всего 2% от общего числа альвеол, поэтому газообмен между содержащимся в дыхательных бронхиолах воздухом и кровью легочных капилляров не может быть значительным. Составляемый этими тремя генерациями, объем равен почти 200 мл. Суммарная площадь поперечного сечения всех бронхиол 19-й генерации составляет 9944 см2, т.е. превышает площадь поперечного сечения трахеи уже в 372 раза.
Последние 4 генерации бронхиол (20, 21, 22, 23) представляют собой альвеолярные ходы и альвеолярные мешочки, которые непосредственно переходят в альвеолы. Суммарный объем альвеолярных ходов и альвеолярных мешочков (без примыкающих к ним альвеол) составляет свыше 1300 мл, а суммарная площадь поперечного сечения альвеолярных мешочков достигает огромной величины — 11800 см2.
Вентиляция легких. Воздухопроводящие пути, легочная паренхима, плевра, костно-мышечный каркас грудной клетки и диафрагма составляют единый рабочий орган, посредством которого осуществляется вентиляция легких.
345

 Вентиляцией легких называют процесс обновления газового состава альвеолярного воздуха, обеспечивающего поступление в них кислорода и выведение избыточного количества углекислого газа. Интенсивность вентиляции определяется глубиной вдоха и частотой дыхания. Наиболее информативным показателем вентиляции легких служит минутный объем дыхания, определяемый как произведение дыхательного объема на число дыханий в минуту. У взрослого мужчины в спокойном состоянии минутный объем дыхания составляет 6- 10 л/мин, при работе — от 30 до 100 л/мин. Частота дыхательных движения в покое 12-16 в 1 мин. Для оценки потенциальных возможностей спортсменов и лиц специальных профессий используют пробу с произвольной максимальной вентиляцией легких, которая у этих людей может достигать 180 л/мин.
Вентиляцией легких называют процесс обновления газового состава альвеолярного воздуха, обеспечивающего поступление в них кислорода и выведение избыточного количества углекислого газа. Интенсивность вентиляции определяется глубиной вдоха и частотой дыхания. Наиболее информативным показателем вентиляции легких служит минутный объем дыхания, определяемый как произведение дыхательного объема на число дыханий в минуту. У взрослого мужчины в спокойном состоянии минутный объем дыхания составляет 6- 10 л/мин, при работе — от 30 до 100 л/мин. Частота дыхательных движения в покое 12-16 в 1 мин. Для оценки потенциальных возможностей спортсменов и лиц специальных профессий используют пробу с произвольной максимальной вентиляцией легких, которая у этих людей может достигать 180 л/мин.
Разные отделы легких человека вентилируются неодинаково, в зависимости от положения тела. При вертикальном положении человека нижние отделы легких вентилируются лучше, чем верхние. Если человек лежит на спине, то разница в вентиляции верхушечных и нижних отделов легких исчезает, однако, при этом задние (дорсальные) их участки начинают вентилироваться лучше, чем передние (вентральные). В положении лежа на боку лучше вентилируется легкое, находящееся снизу. Неравномерность вентиляции верхних и нижних участков легкого при вертикальном положении человека связана с тем, что транспульмональное давление (разность давления в легких и плевральной полости) как сила, определяющая объем легких и его изменения, у этих участков легкого не одинаково. Поскольку легкие обладают весом, у их основании транспульмональное давление меньше, чем у верхушек. В связи с этим нижние отделы легких в конце спокойного выдоха более сдавлены, однако, при вдохе они расправляются лучше, чем верхушки. Этим объясняется и более интенсивная вентиляция отделов легких, оказавшихся снизу, если человек лежит на спине или на боку.
В конце выдоха объем газов в легких равен сумме остаточного объема и резервного объема выдоха, т.е. представляет собой так называемую функциональную остаточную емкость легких (ФОЕ). В конце вдоха этот объем увеличивается на величину дыхательного объема, т.е. того объема воздуха, который поступает в легкие во время вдоха и удаляется из них во время выдоха.
Поступающий в легкие во время вдоха воздух заполняет дыхательные пути, и часть его достигает альвеол, где смешивается с альвеолярным воздухом. Остальная, обычно меньшая, часть остается в дыхательных путях, в которых обмен газов между содержащимся в них воздухом и кровью не происходит, т.е. в так называемом мертвом пространстве.
Дыхательное мертвое пространство — объем дыхательных путей, в котором не происходят процессы газообмена между воздухом и кровью. Различают анатомическое и физиологическое (или функциональное) мертвое пространство. Анатомическое дыхательное мертвое пространство представляет собой объем воздухоносных путей, начиная от отверстий носа и рта и кончая дыхательными бронхи-
346


 олами легкого. Под функциональным (физиологическим) мертвым пространством понимают все те участки дыхательной системы, в которых не происходит газообмена. К функциональному мертвому пространству в отличие от анатомического относятся не только воздухоносные пути, но также альвеолы, которые вентилируются, но не перфузируются кровью. В таких альвеолах газообмен невозможен, хотя их вентиляция и происходит.
олами легкого. Под функциональным (физиологическим) мертвым пространством понимают все те участки дыхательной системы, в которых не происходит газообмена. К функциональному мертвому пространству в отличие от анатомического относятся не только воздухоносные пути, но также альвеолы, которые вентилируются, но не перфузируются кровью. В таких альвеолах газообмен невозможен, хотя их вентиляция и происходит.
У человека среднего возраста объем анатомического мертвого пространства равен 140-150 мл или примерно 1/3 дыхательного объема при спокойном дыхании. В альвеолах к концу спокойного выдоха находится около 2500 мл воздуха (функциональная остаточная емкость), поэтому при каждом спокойном вдохе обновляется лишь 1/7 часть альвеолярного воздуха.
Таким образом, вентиляция обеспечивает поступление наружного воздуха в легкие и части его в альвеолы и удаление вместо него смеси газов (выдыхаемого воздуха), состоящей из альвеолярного воздуха и той части наружного воздуха, которая заполняет мертвое пространство в конце вдоха и удаляется первой в начале выдоха. Поскольку альвеолярный воздух содержит меньше кислорода и больше углекислого газа, чем наружный, суть вентиляции легких сводится к доставке в альвеолы кислорода (возмещающего убыль кислорода, переходящего из альвеол в кровь легочных капилляров) и удалению из них углекислого газа (поступающего в альвеолы из крови легочных капилляров). Между уровнем тканевого метаболизма (скорость потребления тканями кислорода и образования в них углекислоты) и вентиляцией легких существует зависимость, близкая к прямой пропорциональности. Соответствие легочной и, главное, альвеолярной вентиляции уровню метаболизма обеспечивается системой регуляции внешнего дыхания и проявляется в виде увеличения минутного объема дыхания (как за счет увеличения дыхательного объема, так и частоты дыхания) при увеличении скорости потребления кислорода и образования углекислоты в тканях.
Вентиляция легких происходит, благодаря активному физиологическому процессу (дыхательным движениям), который обуславливает механическое перемещение воздушных масс по трахеобронхиальным путям объемными потоками. В отличие от конвективного перемещения газов из окружающей среды в бронхиальное пространство дальнейший транспорт газов (переход кислорода из бронхиол в альвеолы и, соответственно, углекислого газа из альвеол в бронхиолы) осуществляется, главным образом, путем диффузии. Поэтому различают понятие "легочная вентиляция" и "альвеолярная вентиляция".
Альвеолярную вентиляцию не удается объяснить только за счет создаваемых активным вдохом конвективных потоков воздуха в легких. Суммарный объем трахеи и первых 16 генераций бронхов и бронхиол составляет 175 мл, последующих трех (17-19) генераций бронхиол — еще 200 мл. Если все это пространство, в котором почти отсутствует газообмен, "промывалось" бы конвективными потоками наружного воздуха, то дыхательное мертвое пространство должно было бы составлять почти 400 мл. Если вдыхаемый воздух
347


 поступает в альвеолы через альвеолярные ходы и мешочки (объем которых равен 1300 мл) также путем конвективных потоков, то кислород атмосферного воздуха может достигнуть альвеол лишь при объеме вдоха не менее 1500 мл, тогда как обычный дыхательный объем составляет у человека 400- 500 мл.
поступает в альвеолы через альвеолярные ходы и мешочки (объем которых равен 1300 мл) также путем конвективных потоков, то кислород атмосферного воздуха может достигнуть альвеол лишь при объеме вдоха не менее 1500 мл, тогда как обычный дыхательный объем составляет у человека 400- 500 мл.
В условиях спокойного дыхания (частота дыхания 15 а мин, продолжительность вдоха 2 с, средняя объемная скорость вдоха 250 мл/с), во время вдоха (дыхательный объем 500 мл) наружный воздух заполняет всю проводящую (объем 175 мл) и переходную (объем 200 мл) зоны бронхиального дерева. Лишь небольшая его часть (менее 1/3) поступает в альвеолярные ходы, объем которых в несколько раз превышает эту часть дыхательного объема. При таком вдохе линейная скорость потока вдыхаемого воздуха в трахее и главных бронхах равна примерно 100 см/с. В связи с последовательным делением бронхов на все более меньшие по диаметру, при одновременном увеличении их числа и суммарного просвета каждой последующей генерации, движение по ним вдыхаемого воздуха замедляется. На границе проводящей и переходной зон трахеобронхиального пути линейная скорость потока составляет всего около 1 см/с, в дыхательных бронхиолах она снижается до 0.2 см/с, а в альвеолярных ходах и мешочках — до 0.02 см/с.
Таким образом, скорость конвективных потоков воздуха, возникающих во время активного вдоха и обусловленных разностью между давлением воздуха в окружающей среде и давлением в альвеолах.в дистальных отделах трахеобронхиального дерева весьма мала, а в альвеолы из альвеолярных ходов и альвеолярных мешочков воздух поступает путем конвекции с небольшой линейной скоростью. Однако, суммарная площадь поперечного сечения не только альвеолярных ходов (тысячи см2), но и дыхательных бронхиол, образующих переходную зону (сотни см2), достаточно велика для того, чтобы обеспечить диффузионный перенос кислорода из дистальных отделов бронхиального дерева в альвеолы, а углекислого газа — в обратном направлении. Благодаря диффузии, состав воздуха в воздухоносных путях респираторной и переходной зоны приближается по составу к альвеолярному. Следовательно, диффузионное перемещение газов увеличивает объем альвеолярного и уменьшает объем мертвого пространства. Кроме большой площади диффузии, этот процесс обеспечивается также значительным градиентом парциальных давлений: во вдыхаемом воздухе парциальное давление кислорода на 6.7 кПа (50 мм рт.ст.) больше, чем в альвеолах, а парциальное давление углекислого газа в альвеолах на 5.3 кПа (40 мм рт.ст.) больше, чем во вдыхаемом воздухе. В течение одной секунды за счет диффузии концентрация кислорода и углекислоты в альвеолах и ближайших структурах (альвеолярные мешочки и альвеолярные ходы) практически выравниваются.
Следовательно, начиная с 20-й генерации, альвеолярная вентиляция обеспечивается исключительно за счет диффузии. Благодаря диффузионному механизму перемещения кислорода и углекислого газа, в легких отсутствует постоянная граница между мертвым про-
348


 странством и альвеолярным пространством. В воздухоносных путях есть зона, в пределах которой происходит процесс диффузии, где парциальное давление кислорода и углекислого газа изменяется, соответственно, от 20 кПа (150 мм рт.ст.) и 0 кПа в проксимальной части бронхиального дерева до 13.3 кПа (100 мм рт.ст.) и 5.3 кПа (40 мм рт.ст.) в дистальной его части. Таким образом, по ходу бронхиальных путей существует послойная неравномерность состава воздуха от атмосферного до альвеолярного (рис.8.4). Эта зона смещается в зависимости от режима дыхания и, в первую очередь, от скорости вдоха; чем больше скорость вдоха (т.е. в итоге, чем больше минутный объем дыхания), тем дистальнее по ходу бронхиального дерева выражены конвективные потоки со скоростью, превалирующей над скоростью диффузии. В результате с увеличением минутного объема дыхания увеличивается мертвое пространство, а граница между мертвым пространством и альвеолярным пространством сдвигается в дистальном направлении. Следовательно, анатомическое мертвое пространство (если его определять числом генераций бронхиального дерева, в которых диффузия еще не имеет значения) изменяется так же, как и функциональное мертвое пространство — в зависимости от объема дыхания.
странством и альвеолярным пространством. В воздухоносных путях есть зона, в пределах которой происходит процесс диффузии, где парциальное давление кислорода и углекислого газа изменяется, соответственно, от 20 кПа (150 мм рт.ст.) и 0 кПа в проксимальной части бронхиального дерева до 13.3 кПа (100 мм рт.ст.) и 5.3 кПа (40 мм рт.ст.) в дистальной его части. Таким образом, по ходу бронхиальных путей существует послойная неравномерность состава воздуха от атмосферного до альвеолярного (рис.8.4). Эта зона смещается в зависимости от режима дыхания и, в первую очередь, от скорости вдоха; чем больше скорость вдоха (т.е. в итоге, чем больше минутный объем дыхания), тем дистальнее по ходу бронхиального дерева выражены конвективные потоки со скоростью, превалирующей над скоростью диффузии. В результате с увеличением минутного объема дыхания увеличивается мертвое пространство, а граница между мертвым пространством и альвеолярным пространством сдвигается в дистальном направлении. Следовательно, анатомическое мертвое пространство (если его определять числом генераций бронхиального дерева, в которых диффузия еще не имеет значения) изменяется так же, как и функциональное мертвое пространство — в зависимости от объема дыхания.
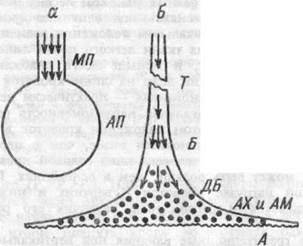
Рис.8.4. Схема альвеолярной вентиляции.
"а" — по устаревшим и "б" — по современным представлениям. МП — мертвое пространство; АП — альвеолярное пространство; Т — трахея; Б — бронхи; ДБ — дыхательные бронхиолы; АХ — альвеолярные ходы; AM — альвеолярные мешочки; А — альвеолы. Стрелками обозначены конвективные потоки воздуха, точками — область диффузионного обмена газов.
 Соотношение вентиляции и перфузии легких.Количество альвеол в одном легком человека равно приблизительно 300 млн. Суммарная площадь альвеоло-капиллярного барьера, через который происходит обмен газами между альвеолярным воздухом и смешанной
Соотношение вентиляции и перфузии легких.Количество альвеол в одном легком человека равно приблизительно 300 млн. Суммарная площадь альвеоло-капиллярного барьера, через который происходит обмен газами между альвеолярным воздухом и смешанной
349



 венозной кровью, имеет огромные размеры (70-80 м2). Это достигается за счет большой суммарной площади альвеол и необычайной плотности расположения легочных капилляров, сеть которых образует как бы почти сплошной слой крови на поверхности альвеол. Этот слой является столь тонким, что объем крови в легочных капиллярах, несмотря на значительную его поверхность, составляет всего 100-150 мл из общего количества 500-600 мл крови, одновременно содержащейся в малом круге кровообращения. Большая площадь альвеоло-капиллярного барьера и его минимальная толщина (от 0.3 до 2.0 мкм) создают оптимальные условия для диффузии кислорода из альвеолярного воздуха в кровь легочных капилляров, а углекислого газа — в противоположном направлении. Средняя продолжительность пребывания эритроцитов в легочных капиллярах составляет, в зависимости от скорости легочного кровотока, 0.25-0.75 с, что достаточно для того, чтобы оксигенация крови практически успела закончиться даже при поступлении к легким венозной крови с очень низким содержанием кислорода.
венозной кровью, имеет огромные размеры (70-80 м2). Это достигается за счет большой суммарной площади альвеол и необычайной плотности расположения легочных капилляров, сеть которых образует как бы почти сплошной слой крови на поверхности альвеол. Этот слой является столь тонким, что объем крови в легочных капиллярах, несмотря на значительную его поверхность, составляет всего 100-150 мл из общего количества 500-600 мл крови, одновременно содержащейся в малом круге кровообращения. Большая площадь альвеоло-капиллярного барьера и его минимальная толщина (от 0.3 до 2.0 мкм) создают оптимальные условия для диффузии кислорода из альвеолярного воздуха в кровь легочных капилляров, а углекислого газа — в противоположном направлении. Средняя продолжительность пребывания эритроцитов в легочных капиллярах составляет, в зависимости от скорости легочного кровотока, 0.25-0.75 с, что достаточно для того, чтобы оксигенация крови практически успела закончиться даже при поступлении к легким венозной крови с очень низким содержанием кислорода.
Для полного насыщения крови кислородом в легочных капиллярах
необходимо, чтобы кровоток во всех участках легких оптимально
соответствовал вентиляции этих участков. Однако, распределение
кровотока по легким у человека, как оказалось, не является равно
мерным, и кровоснабжение разных участком легких зависит от по
ложения тела человека, изменяясь под влиянием гравитационного
фактора. У человека в вертикальном положении величина легочного
кровотока на единицу объема ткани легкого почти линейно убывает
в направлении снизу вверх, и меньше всего снабжаются кровью
верхушки легких. В положении лежа на спине кровоток в верхушках
легких увеличивается, а в основаниях — практически не изменяется,
в результате чего, его вертикальная неравномерность распределения
почти исчезает. Однако, в этом положении кровоток в задних (до
рсальных) отделах легких становится выше, чем в передних (вен
тральных). При положении человека вниз головой кровоток в вер
хушках легких может быть больше, чем в основаниях. При умерен
ной физической нагрузке кровоток в верхних и нижних отделах
легких увеличивается и регионарные различия его распределения
сглаживаются.
Дата добавления: 2016-03-27; просмотров: 2477;