МЕХАНИЗМЫ ЗАМЫКАНИЯ ВРЕМЕННОЙ СВЯЗИ
До настоящего времени нет единой точки зрения на механизмы замыкания временных связей между нейронами высших отделов ЦНС в процессе образования условных рефлексов. Морфологическое объяснение, выдвинутое еще в прошлом веке, предполагало, что во время образования условного рефлекса происходит рост отростков нервных клеток, устанавливающих новые межклеточные связи. Сходные идеи высказывались и позже, в частности относительно роли своеобразных выростов — «шипиков» на апикальных дендритах пирамидных нейронов.
Функциональное объяснение видело ответ на поставленный вопрос в повышенной проводимости существующих синапсов, которая может обусловливаться непрерывной циркуляцией по нейронным кругам, увеличением числа используемых синапсов, т.е. явлениями, подобными посттетанической потенциации.
Было высказано предположение и о возможной роли глин в организации условно-рефлекторной деятельности. В настоящее время накопилось много фактов, свидетельствующих о том, что функция глиальной ткани не сводится лишь к опорной и трофической. Возможность участия глиальных клеток в образовании стойкой временной связи заключается в миелинизации «голых» пресинаптических терминалей аксонов, что делает их способными быстро проводить импульсы.
Многие исследователи считают ведущим фактором изменения синтеза белка в клетках. Исследование нуклеотидного состава рибонуклеиновой кислоты (РНК) в процессе выработки условных рефлексов позволило предположить, что совместное возбуждение многих нейронов отражается на структуре РНК, в которой кодируется сигнальное раздражение. В свете этой гипотезы трактовались результаты экспериментов, показавших облегчение выработки новых навыков у червей, которым скормили тех, у кого этот навык был заранее выработан. Такой «перенос химической памяти» был описан рядом исследователей на основе опытов с введением различным животным экстрактов из мозга животных, обученных какому-либо навыку и его устранение при действии фермента рибонуклеазы, разрушающей РНК (ряд исследователей не подтвердили эти результаты). Некоторые считают более вероятным, что след памяти закрепляется на молекулах дезоксирибонуклеиновой кислоты (ДНК), которая сохраняет этот след дольше, чем относительно быстро синтезируемая РНК. Изменение свойств ДНК и РНК реализуется через количественные и качественные изменения синтеза белков. Описаны специфические белковые вещества мозга, с которыми связывают фиксацию временной связи между нейронами вплоть до попыток выделения полипептидных комплексов, ответственных за воспроизведение оборонительных, пищевых и других условных рефлексов.
В других гипотезах на первый план в механизмах формирования временной связи выдвигаются системы белков-ферментов и их ингибиторов, которые могут изменять свое состояние.
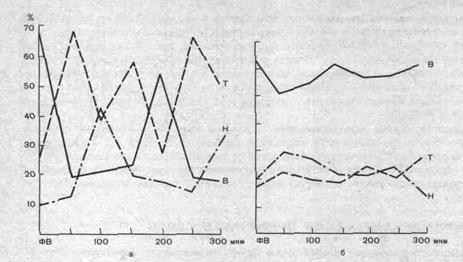
Рис. 236. Распределение возбуждающихся (В), тормозящихся (Т) и нереагирующих (Н) нейронов зрительной коры мозга морской свинки в фокусе возбуждения (ФВ) и на разном удалении от него (по А. Б. Когану и Подладчиковой).
а — первичные реакции 502 нейронов; б — вторичные реакции 493 нейронов. На ординате — процентные доли нейронов В, Т, Н; на абсциссе — расстояние от фокуса возбуждения.
Переход от изучения деятельности отдельных нейронов к изучению их взаимодей
ствия приводит к трактовке механизма временной связи условного рефлекса как форми
рования определенного узора нейронной активности на основе избирательной общности
свойств реагирования. Возникновение узора определяется такими условиями, которые
облегчают проведение возбуждения в структурах мозга: синхронность и синфазность
активности в определенном диапазоне частот (переход значительного числа нейронов,
образующих цепочки возбуждения, на сходные уровни лабильности). Взаимодействие
очагов возбуждения, ограниченных торможением, которое формируется под влиянием
афферентных сигналов, имеет динамический характер и наиболее выражено в фазу пер
вичной активации нейронов (рис. 236). За последнее время представление о временной
связи как пространственно-временной организации межнейронных отношений (получив
шей название нейроселективного действия) подкрепляется и нейрохимическими иссле
дованиями, которые позволили сделать заключение о специфическом изменении химизма
нейронов, объединяющем их в функциональные ансамбли, об избирательной активации
через ДНК нейроглиальных комплексов. х
ПАМЯТЬ
Временная нервная связь, лежащая в основе образования условного рефлекса, — лишь частный случай общебиологического свойства хранения воспринятой информации, которое в той или иной степени проявляется у всех живых существ, начиная с простейших.
Временную связь оказалось возможным выработать у растений, например, на базе защитной реакции стыдливой мимозы или суточного цикла складывания листьев у бобовых.
В широком смысле такое свойство, обеспечивающее запечатлевание связей событий окружающего мира, что позволяет накапливать и использовать жизненный опыт, является памятью. О механизме возникающих при этом межклеточных временных связей пока мало известно.
|
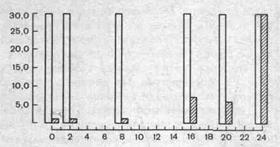
Рис. 237. Определение времени консолидации следа долгосрочной памяти при образовании оборонительного условного рефлекса (задержки выхода мыши на платформу), за которым следовал удар тока.
На ординате — задержка выхода на платформу в секундах, на абсциссе — время в минутах от сочетания выхода с ударом тока до воздействия кратковременным эфирным наркозом. Белые столбики — выработка рефлекса без наркотизации, заштрихованные — после наркотизации в разные сроки.
В нервной системе механизмы памяти получили чрезвычайное развитие и приобрели ведущую роль в поведении. По проявлениям различают память образную, которая воспроизводит образ жизненно важного объекта, эмоциональную, когда аналогичная ситуация вызывает эмоции, сопровождавшие происходившие ранее в этой ситуации события, и словесно-логическую, свойственную только человеку (И. С. Беритов). По времени сохранения различают кратковременную и долговременную память, имеющие в своей основе разные механизмы. Кратковременную память называют также оперативной, так как она обеспечивает выполнение текущих операций мышления (например, умножая 26 на 4, пишем 4, а 2 «держим в уме», чтобы через несколько секунд прибавить к 8). 'Предполагают, что механизмом кратковременной памяти может быть циркуляция импульсных потоков по замкнутым кругам нейронных цепей. Этому соответствует то обстоятельство, что кратковременная память легко нарушается под действием сильных посторонних раздражений, наркозом, электрической стимуляцией мозга, его гипоксией, различными нейротропными ядами, которые при той же интенсивности действия не затрагивают память долговременную. В основе же долговременной памяти лежат, по-видимому, сложные процессы, связанные с активностью синтеза белковых молекул в клетках большого мозга.
Как показано на рис. 237, время, необходимое для перехода кратковременной памяти в долговременную, в условиях эксперимента составляет около 30 с. Однако существует мнение, что речь идет лишь о последовательных этапах консолидации единого механизма памяти, а не о двух ее разновидностях. Сопоставление обширного материала клинических наблюдений и результатов разнообразных экспериментов на животных позволяет считать, что память как единый процесс складывается по крайней мере из трех взаимосвязанных этапов, возможно, имеющих различные механизмы: запоминания, хранения опыта и воспоминания (воспроизведение опыта).
Дата добавления: 2016-03-27; просмотров: 1383;