ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань очень широко распространена в организме: она входит в состав стенки полых (трубчатых) внутренних органов - бронхов, желудка, кишки, матки, маточных труб, мочеточников, мочевого пузыря (висцеральная гладкая мышечная ткань), а также сосудов (васкулярная гладкая мышечная ткань). Васкулярная гладкая мышечная ткань отличается от висцеральной некоторыми структурными, биохимическими и функциональными особенностями, чувствительностью к действию ряда гормонов, нейромедиаторов и фармакологических препаратов. Гладкая мышечная ткань встречается также в коже, где она образует мышцы, поднимающие волос, а также в капсулах и трабекулах некоторых органов (селезенка, яичко) (рис. 7.16).
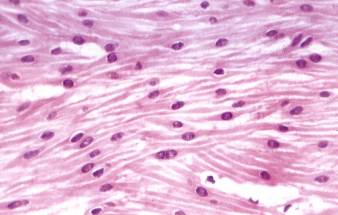
Рис. 7.16. На микрофотографии (х480)изображен продольный срез гладкой мышечной ткани. Мышечная клетка имеет веретенообразную форму, в центре расположено ядро. Концы клеток вклиниваются между срединными частями соседних клеток, благодаря чему создается связь клеток в пласте. Между гладкомышечными клетками располагается соединительная ткань.
Движения, осуществляемые гладкой мышечной тканью, – сравнительно медленные и продолжительные, она обеспечивает также длительные тонические сокращения. Ее сокращения вызывают изменения величины просвета трубчатых органов и лежат в основе их перистальтики. Благодаря сократительной активности этой ткани обеспечивается деятельность органов пищеварительного тракта, регуляция дыхания, крово- и лимфотока, выделение мочи, транспорт половых клеток и др.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
Структурно-функциональной единицей гладкой мышечной ткани мезенхимного типа служит гладкий миоцит (гладкая мышечная клетка) (рис. 7.16; 7.17).
ГЛАДКИЕ МИОЦИТЫ
Гладкие миоциты - одноядерные клетки преимущественно веретеновидной формы, не обладающие поперечной исчерченностью и образующие многочисленные соединения друг с другом (рис. 7.16, 7.17). Длина клеток в состоянии расслабления варьирует в пределах 20-1000 мкм (составляя, в среднем, около 200 мкм), их толщина колеблется от 2 до 20 мкм. При резком сокращении длина миоцитов может уменьшаться до 20% начальной. Наиболее крупные клетки характерны для стенки внутренних органов (максимальной длины 500-1000 мкм достигают миоциты матки при беременности), самые мелкие (длиной около 20 мкм) располагаются в стенке сосудов. Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной, содержат одно ядро и саркоплазму, в которой располагаются органеллы и включения.
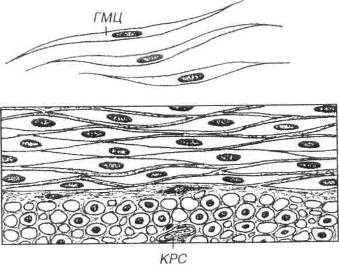
Рис. 7.17. Гладкая мышечная ткань. Сверху показаны изолированные гладкие миоциты (ГМЦ), внизу - их пласт, образованный двумя слоями, в которых клетки ориентированы во взаимно перпендикулярных плоскостях (ГМЦ видны на продольном и поперечном разрезах). КРС - кровеносный сосуд в прослойке рыхлой волокнистой соединительной ткани.
Саркоплазма гладких миоцитов содержит умеренно развитые органеллы общего значения, которые располагаются вместе с включениями в конусовидных участках у полюсов ядра. Периферическая ее часть занята миофиламентами. В саркоплазме выделяют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппарат гладких миоцитовпредставлен тонкими (актиновыми) и толстыми (миозиновыми) филаментами, которые, однако, в отличие от поперечнополосатых мышечных тканей, не формируют миофибрилл.
Тонкие (актиновые) миофиламенты образованы особым набором изоформ актина, свойственным гладким миоцитам, причем помимо мышечного актина в них обнаруживается и немышечный (цитоплазмати-ческий) актин. Тонкие филаменты преобладают над толстыми по количеству и занимаемому объему. Они более многочисленны, чем в поперечнополосатых мышечных тканях и располагаются в саркоплазме пучками по 10-20 филаментов, лежащими параллельно или под углом к длинной оси клетки и образующими сетевидные структуры.
 Толстые (миозиновые филаменты), в отличие от таковых в поперечнополосатой мышечной ткани, обладают различной длиной (при этом они значительно короче тонких нитей), менее стабильны, не содержат центральной гладкой части, поскольку покрыты миозиновыми головками по всей длине. Это обеспечивает более значительное перекрытие тонких и толстых филаментов, а, следовательно, и большую силу сокращения. Относительное содержание миозиновых филаментов в гладких миоцитах ниже, чем в миофибриллах поперечнополосатой мышечной ткани; на один миозиновый филамент в гладких миоцитах приходится не менее 12 актиновых. По мнению некоторых авторов, миозиновые филаменты гладких миоцитов обладают значительной лабильностью и окончательно собираются непосредственно перед сокращением, распадаясь после него.
Толстые (миозиновые филаменты), в отличие от таковых в поперечнополосатой мышечной ткани, обладают различной длиной (при этом они значительно короче тонких нитей), менее стабильны, не содержат центральной гладкой части, поскольку покрыты миозиновыми головками по всей длине. Это обеспечивает более значительное перекрытие тонких и толстых филаментов, а, следовательно, и большую силу сокращения. Относительное содержание миозиновых филаментов в гладких миоцитах ниже, чем в миофибриллах поперечнополосатой мышечной ткани; на один миозиновый филамент в гладких миоцитах приходится не менее 12 актиновых. По мнению некоторых авторов, миозиновые филаменты гладких миоцитов обладают значительной лабильностью и окончательно собираются непосредственно перед сокращением, распадаясь после него.
Сокращение гладких миоцитов обеспечивается взаимодействием актиновых и миозиновых миофиламентов и развивается в соответствии с моделью скользящих нитей. Оно происходит более медленно и длится дольше, чем в скелетной мышце, что обусловлено более низкой скоростью гидролиза АТФ в гладких миоцитах.
Роль Са2+ в сокращении гладких миоцитов. Как и в поперечнополосатых мышечных тканях, сокращение гладких миоцитов индуцируется притоком Са2+ в саркоплазму, который в этих клетках выделяется саркоплазматшеской сетью и кавеолами (см. ниже), а также вследствие увеличения проницаемости сарколеммы для данных ионов.
Образование мостиков типа "щеколды" (latch-bridges в англоязычной литературе) является особенностью сократительного аппарата гладких миоцитов: часть миозиновых мостиков после дефосфорилирования не отсоединяется от актина, а остается с ним связанной. Благодаря этому гладкая мышца способна обеспечивать длительное поддержание тонуса без существенных дополнительных энергетических затрат (так как указанные мостики обладают очень медленной циклической активностью).
Опорный аппарат гладкого миоцитапредставлен его сарколеммой, базальной мембраной, системой элементов цитоскелета и связанных с ними плотных телец.
Сарколемма каждого миоцита окружена базальной мембраной, в которую вплетаются тонкие ретикулярные, коллагеновые и эластические волокна; коллагеновые фибриллы, прикрепляющиеся к сарколемме в области ее углублений по краям миоцитов, воспринимают усилие, развивающееся при сокращении клеток.
Плотные тельца, связанные с сарколеммой, некоторые авторы называют плотными пластинками и считают структурами, не идентичными расположенным в саркоплазме (поскольку они различаются по химическому составу). Более того, в соответствии с современными представлениями, плотные пластинки лишь на срезах кажутся отдельными небольшими образованиями, в реальности же они имеют вид длинных непрерывных "ребер", идущих параллельно друг другу по внутренней поверхности сарколеммы вдоль длинной оси миоцита.
Плотные пластинки включают периферический и глубокий слои. Первый прилежит к сарколемме и образован филаментами немышечного актина, связанными с трансмембранными белками интегринами по средством комплекса адгезивных белков (винкулина, талина, тетина и др.). В глубоком слое филаменты мышечного актина прикреплены к молекулам немышечного актина связующими белками (например, филамином).
Аппарат передачи возбуждения(с сарколеммы на сократительный аппарат) в гладких миоцитах изучен недостаточно. К нему относят саркоплазматическую сеть, которая в этих клетках рудиментарна и состоит из системы мелких цистерн и пузырьков, а также особые мембранные структуры – кавеолы. Т-трубочки отсутствуют.
Кавеолы- колбовидные впячивания поверхности сарколеммы диаметром около 70 нм (с более узкой "шейкой"), располженные перпендикулярно длинной оси клетки. Кавеолы открыты в сторону межклеточного пространства, часто располагаются рядами вдоль длинной оси миоцита (занимая промежутки между плотными пластинками), иногда уходят вглубь его саркоплазмы в виде ветвящихся цепочек. Они очень многочисленны (до нескольких сотен тысяч в одной клетке); площадь их суммарной поверхности составляет около 1/3 площади поверхности сарколеммы. Число кавеол не меняется при сокращении, расслаблении или растяжении клетки, они, по-видимому, не участвуют в процессах эндоцитоза. Кавеолы содержат высокие концентрации кальция, а в их мембране имеются белки, обеспечивающие транспорт кальция в саркоплазму и из нее. Местами они контактируют с элементами саркоплазматической сети. Кавеолы, по-видимому, не только гомологичны системе Т-трубочек поперечнополосатых мышечных тканей, но и выполняют ряд функций, свойственных саркоплазматической сети (рис. 7.18).
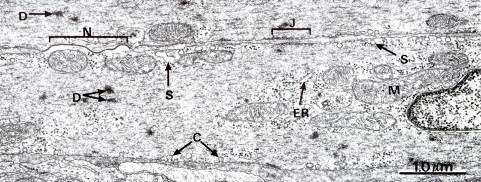
Рис. 7.18. Электроннограмма гладкой мышечной ткани (х21.000). Заметны детали стоения плазматической и внутриклеточной мембран. Плазматическая мембрана имеет внутриклеточные впячивания – кавеолы (С). Внутриклеточные мембраны: – элементы ЭПР (SR), комплекс Гольджи. Везикулярные структуры S, располагаются вблизи плазмолеммы, часто соединяются с кавеолами. Это структуры, аналогичные по-видимому СПР скелетных мышц, N – межклеточные контакты, N – нексус, J – адгезивный контакт, D – электронноплотные гранулы.
Энергетический аппарат гладких миоцитовпредставлен митохондриями, а также включениями гранул гликогена и мелкими липидными каплями преимущественно у полюсов ядра
Синтетический аппарат гладких миоцитовпредставлен элементами грЭПС и комплексом Гольджи, лежащими у полюсов ядра, а также свободными рибосомами, которые располагаются, наряду с этими участками, по всей саркоплазме. Благодаря выраженной синтетической активности гладкие миоциты продуцируют и выделяют (подобно фибробластам) коллагены, эластин и компоненты аморфного вещества, ряд факторов роста и цитокинов.
Лизосомальный аппарат (аппарат внутриклеточного переваривания) гладких миоцитов развит сравнительно слабо.
Дата добавления: 2016-02-16; просмотров: 6826;