Лекция 7. КООРДИНАЦИЯ РЕФЛЕКТОРНЫХ ПРОЦЕССОВ
Рефлексы – это элементы целостного приспособительного поведения организма, имеющий определенный физиологический смысл, и эти элементы взаимодействуют между собой так, что обеспечивают общий полезный для организма результат. Взаимоотношения между рефлексами могут быть содружественными и антагонистическими. Содружественные рефлексы обычно входят в одну функциональную систему, так как имеют общий результат. Например, чтобы двигаться с максимальной скоростью, необходимы не только интенсивная работа скелетных мышц, но и вегетативное обеспечение этой работы – учащение и усиление сердечных сокращений, учащение дыхания. Или пищевые рефлексы – жевание, глотание, отделение пищеварительных соков – обеспечивают поступление в организм питательных веществ. При антагонистических взаимодействиях один рефлекс полностью или частично подавляет другой. Например, рефлексы, возникающие при болевых раздражителях, подавляют рефлексы пищевого поведения, или рефлекторный акт глотания в норме не может осуществляться одновременно с актом вдоха.
Согласованное взаимодействие рефлекторных процессов называется координацией. Остановимся на некоторых принципах координации, которым подчиняются все нервные центры.
Принцип конвергенции и принцип иррадиации возбуждения распространяются не только на работу отдельного нервного центра, но и наблюдается при взаимодействии различных рефлекторных актов. Так, раздражение определенного рецептивного поля может вызвать соответствующий рефлекс, а усиление действия раздражителя приведет к появлению нескольких рефлекторных реакций (иррадиация возбуждения с одного НЦ на другие). Например, свет вызывает сужение зрачка, яркий свет – мигательный рефлекс. Конвергенция при взаимодействии рефлексов лежит в основе принципа «общего конечного пути», согласно которому одна и та же поведенческая реакция может возникать при раздражении различных рецептивных полей. Например, слюноотделение происходит при раздражении вкусовых и тактильных рецепторов полости рта, ориентировочная реакция – при появлении новых зрительных и слуховых раздражителей, сгибание конечности – при ударе по сухожилию и при раздражении болевых рецепторов. Двигательная активность может повышаться при раздражении центров голода гипоталамуса, а также при угрожающих воздействиях. Это объясняется тем, что одни и те же эффекторные нейроны входят в дуги различных рефлексов.
Принцип взаимодействия возбуждения и торможения.Торможение – это уменьшение или прекращение возбуждения. Так же как и возбуждение, торможение имеет определенную силу и длительность. Играет важнейшую роль в координации рефлекторных процессов и является необходимым условием нормальной деятельности НС. Торможение сопутствует возбудительному процессу, препятствует чрезмерному распространению возбуждения. Явление центрального торможения было открыто И.М.Сеченовым в 1862 г. В своей работе «Рефлексы головного мозга» он описывает уменьшение защитных спинальных рефлексов лягушки при наложении на разрез промежуточного мозга лягушки кристалла поваренной соли. Впоследствии оказалось, что это торможение связано с раздражением тормозных структур ретикулярной формации. Значительный вклад в изучение торможения внесли Н.Е. Введенский (пессимальное торможение), Дж. Экклс, И.С. Бериташвили и др.
Торможение может быть первичным и вторичным. Первичное развивается в результате активности тормозных нейронов, использующих тормозные медиаторы. Разновидностями первичного торможения являются возвратное и латеральное. Нейроны, обеспечивающие возвратное торможение, называются клетками Реншоу. Тормозная клетка Реншоу возбуждается коллатералью аксона нейрона при его интенсивном возбуждении, таким образом происходит саморегуляция ритма активности нейронов в различных структурах ЦНС. Латеральное торможение проявляется в том, что коллатерали аксонов возбужденных нейронов активируют тормозные клетки, действующие на нейроны, вызывающие другие реакции, например, глотание во время вдоха или сокращение мышц – сгибателей во время разгибания конечности (рис.9).

| 
|
| ]А | Б |
Рис.9. Разновидности первичного торможения: А – возвратное торможение;
Б – латеральное торможение; НЦ – нервный центр; РО – рабочий орган
Вторичное торможение – развивается в результате его собственного возбуждения, т.е. вторично по отношению к нему. Различают пессимальное и последовательное вторичное торможение. Пессимальное торможение развивается в результате чрезмерной активности нервной клетки, когда она истощается – снижаются энергетические запасы, необходимые для восстановления ионных градиентов. Может возникать как в нервных центрах, так и на уровне отдельных нейронов и синапсов. Такое торможение имеет ярко выраженную охранительную роль. Последовательное торможение формируется вслед за обычным по силе возбуждением, если имеется длительная следовая деполяризация. Позволяет ограничить ритм активности нервной клетки (например, в мотонейронах спинного мозга).
Взаимодействие возбуждения и торможения в нервной системе может проявляться по-разному. Один из вариантов – сопряженное взаимодействие, или одновременная индукция: возбужденный НЦ посредством латерального торможения выключает активность центров, несовместимых с данной реакцией (рис.10).

| Рис.10. Одновременная индукция возбуждения и торможения. НЦ – нервный центр; КР – клетка Реншоу |
Впервые это явление было описано Н.Е.Введенским для спинальных мотонейронов, иннервирующих мышцы-сгибатели и мышцы – разгибатели конечностей. Например, чтобы согнуть руку при болевом раздражении необходимо сокращение мышц–сгибателей и расслабление разгибателей. Таким же образом организованы и более сложные рефлексы – например, пищевые, половые, оборонительные. Реципрокные отношения между различными центрами могут изменяться и подавляться влияниями высших отделов ЦНС. Так, при ходьбе мы, делая шаг левой ногой, выбрасываем вперед правую руку, но мы можем при некотором усилии выбрасывать вперед и одноименную руку. Как только волевой контроль ослабевает, восстанавливается естественная координация движений. Подобный механизм отчасти объясняет всем известную истину, что лучший отдых – это смена деятельности (при умственном утомлении целесообразно привести в активное состояние двигательные центры, при этом активность структур, участвующих в умственной деятельности, затормозится, и отдых будет более полноценным).
Другой вариант взаимодействия возбуждения и торможения – последовательная индукция: в нервных центрах возбуждение и торможение последовательно сменяют друг друга, причем после торможения наблюдается усиление возбуждения (положительная последовательная индукция), а вслед за возбуждением развивается усиленное торможение (отрицательная последовательная индукция). Например, возбуждение мотонейронов мышц – сгибателей после длительного сохранения разгибания будет более сильным. Положительную последовательную индукцию в этом случае можно объяснить тем, что при сгибании конечности центр разгибания находился в состоянии реципрокного торможения, но к нему постоянно поступали импульсы от проприорецепторов растянутой мышцы. К моменту окончания сгибания центр разгибания быстро приходит в возбужденное состояние. Подобный феномен отдачи обеспечивает ритмические рефлексы (чесание, шагание). Такое взаимодействие часто наблюдается в поведении. У детей волевое торможение двигательной активности во время урока приводит к резкому ее повышению во время перемены. Подобное явление наблюдается при разных формах психической деятельности: например, если проявление эмоции долго сдерживается волевым усилием, то при ослаблении контроля чувства «выплескиваются» с удвоенной силой.
Принцип обратной связи. Любой рефлекторный акт возникает в ответ на раздражение рецепторов. Но сама рефлекторная реакция сопровождается возбуждением самых разнообразных рецепторов: при сокращении и расслаблении мышц раздражаются проприорецепторы мышц и сухожилий, при повороте глазных яблок возникает не только проприоцептивное, но и зрительное раздражение. Афферентные импульсы, возникающие в рецепторах в результате любого поведенческого акта, называют вторичными афферентными импульсами, в отличие от первичных – тех, которые вызвали данный поведенческий акт. Значение вторичной афферентации очень велико. Она непрерывно сигнализирует нервным центрам о результате рефлекторной реакции. На основании анализа этой информации включаются следующие реакции, или поведенческий акт изменяется так, чтобы эффект был наилучшим. Такое явление называется обратной связью (ОС). Обратные связи могут быть положительными и отрицательными. При положительной ОС вторичная афферентация усиливает ту реакцию, благодаря которой она возникает. При отрицательной ОС вторичные афферентные импульсы угнетают рефлекторную реакцию. Чаще всего положительные и отрицательные ОС сосуществуют. Благодаря ОС интенсивность возбуждения различных групп нейронов в НЦ и последовательность их включения в реакцию становятся строго согласованными. ОС – важнейшее звено функциональных систем регуляции функций с точки зрения теории функциональных систем, созданной выдающимся русским физиологом П.К.Анохиным.
Принцип доминанты – один из важнейших принципов работы НЦ. Этот принцип был сформулирован А.А. Ухтомским в начале 20 века. Суть его заключается в том, что в естественных условиях в ЦНС периодически возникают доминантные (господствующие) очаги возбуждения, подчиняющие себе работу всех других НЦ. Например, электрическое раздражение участка КБП у собаки вызывает сгибание лапы, но если этот участок раздражать у голодной собаки, то ожидаемой реакции не возникнет, а появятся глотательные движения. По определению Ухтомского, доминанта – это временно господствующая рефлекторная система, направляющая текущую деятельность НЦ, или «рефлекторное внимание». В естественных условиях доминантный очаг возникает под действием внешних или внутренних стимулов, имеющих большое биологическое значение (пищевая, половая, оборонительная доминанты). Доминантный очаг можно создать искусственно – например, воздействуя в эксперименте постоянным током на участок мозга. Основные свойства доминантного очага: повышенная возбудимость, стойкость и инертность возбуждения, которое сохраняется после окончания стимуляции, высокая способность к суммации возбуждения, способность поддерживать возбуждение за счет стимулов, адресованных к другим, не доминантным, нервным центрам, тормозящее влияние на другие нервные центры (рис.11).
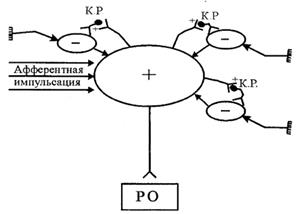
| Рис.11. Тормозящее влияние доминантного очага на други нервные центры. КР – клетка Реншоу; РО – рабочий орган |
Принцип доминанты имеет очень важное адаптивное значение – он позволяет организму реализовывать ту деятельность, которая в данный момент является наиболее важной. Формирование патологического доминантного очага может лежать в основе многих заболеваний – например, неврозов, гипертонической болезни. Яркий пример патологической доминанты – очаг судорожной готовности при эпилепсии. Причиной прекращения доминантного возбуждения может быть удовлетворение потребности, вызвавшей формирование доминанты, запредельное торможение, появление новой доминанты, волевое торможение. Личностная зрелость человека, по мнению Ухтомского, проявляется, в частности, в способности контролировать свои доминанты. Ухтомский говорил, что человек должен стремиться быть командиром, а не жертвой своих доминант.
Принцип субординации нервных центров – проявляется в виде регулирующего влияния вышележащих НЦ на нижележащие. Яркий пример – сеченовское торможение, когда раздражение среднего мозга вызывало торможение спинальных мотонейронов. У высших животных и человека НЦ на всех этажах ЦНС испытывают регулирующие влияния со стороны КБП, что делает возможным волевой контроль движений, эмоциональных реакций и даже, при специальной тренировке, вегетативных функций.
Лекция 6. ПРИНЦИПЫ НЕРВНОЙ РЕГУЛЯЦИИ МОТОРНЫХ ФУНКЦИЙ. ФИЗИОЛОГИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ
Моторные функции обеспечивают поддержание позы и равновесия, а также перемещение тела в пространстве и движения различных частей тела относительно друг друга. Недостаточная мышечная активность, свойственная образу жизни современного человека, нарушает естественные процессы жизнедеятельности, приводя к развитию нездорового состояния организма – гиподинамии. Гиподинамия является фактором риска очень многих заболеваний.
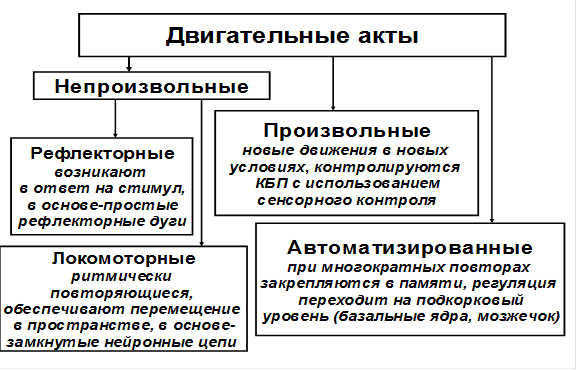
Рабочим органом движения являются мышцы, которые получают сигналы к сокращению от нервной системы.Структуры нервной системы, обеспечивающие регуляцию движений, называются двигательными центрами (ДЦ). ДЦ находятся на всех «этажах» ЦНС, от спинного мозга до коры больших полушарий. Их работа иерархически организована, что является результатом эволюции, в процессе которой происходило приспособление двигательной системы к выполнению все более сложных задач. Наряду с усложнением всей двигательной системы возникала специализация отдельных двигательных центров. ДЦ, непосредственно связанные с мышцами, называются сегментарными. Это скопления мотонейронов в спинном мозге и двигательных ядрах черепно-мозговых нервов ствола головного мозга. ДЦ, регулирующие активность сегментарных центров – надсегментарные ДЦ. На схеме представлена упрощенная классификация движений.
Любой произвольный двигательный акт включает фазу подготовки и фазу реализации движения (рис.12).
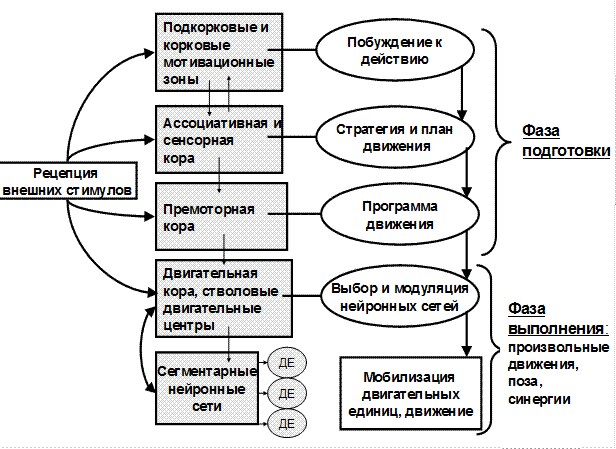
Рис. 12. Схема иерархической организации и системы регуляции произвольных движений
Фаза подготовки начинается с формирования внутреннего побуждения к действию. Формируется двигательная мотивация – стремление к удовлетворению какой-либо доминирующей потребности: пищевой, оборонительной, половой, творческой и др. Нейрофизиологическим субстратом этого этапа являются подкорковые и корковые мотивационные зоны. Далее формируется стратегия и план движения – определяется последовательность движений, их направленность, цель с учетом текущей ситуации (внешние обстоятельства, положение тела и т.д.). Данный этап связан с активностью сенсорных и ассоциативных зон коры. Затем выбирается программа движения (определяется его тактика) – последовательность сокращений и расслаблений определенных мышечных групп. Двигательные программы могут быть врожденными и приобретенными. Первые – это программы ползания, ходьбы, бега, они реализуются у человека не сразу после рождения, а по мере созревания двигательных центров. Вторые - приобретенные программы речи, письма, трудовых и спортивных движений, сформировавшиеся из готовых врожденных «блоков» в результате научения). Выбор двигательной программы – это функция премоторной коры совместно с другими надсегментарными ДЦ (мозжечок, базальные ганглии), которая осуществляется на основе предыдущего этапа и анализа информации об окружающей среде и состоянии опорно-двигательного аппарата.
Фаза реализации (выполнения) движения начинается с модуляции активности сегментарных ДЦ, активации определенных нейронных сетей, что является следствием активности моторной коры и других надсегментарных ДЦ. Затем мобилизуются определенные двигательные единицы сегментарных ДЦ и происходят мышечные сокращения, обеспечивающие целенаправленные движения и поддержание позы. Двигательная единица (ДЕ) – это группа мышечных волокон, иннервируемых одним мотонейроном и сокращающихся одновременно. Наиболее мелкие ДЕ в мышцах, обеспечивающих мелкие дифференцированные движения, а крупные - в мышцах, способных к сильным сокращениям (например, ДЕ наружной прямой мышцы глаза содержит всего 13 волокон, а в ДЕ двуглавой мышцы плеча их 750).
Моторные функции обеспечиваются двумя видами сокращений мышц: фазическими (относительно кратковременными и интенсивными), обеспечивающими движения, и тоническими (длительными сокращениями небольшой интенсивности), обеспечивающими сохранение позы и равновесия. Как правило, активация движений сопровождается снижением тонуса мышц, и наоборот. Между центрами регуляции тонуса и движений существуют реципрокные взаимоотношения. Тем не менее, любое движение требует создания для него удобной позы и адекватного положения тела в пространстве. Сочетание фазных сокращений одних мышц и тонических - других обеспечивает гармонию движений. Принципы координации, которым подчиняются двигательные центры, были рассмотрены в предыдущей лекции.
Дата добавления: 2016-02-13; просмотров: 2782;