Гомеостаз популяций
В природе численность популяций испытывает колебания. В связи с размерами ареала популяций может значительно изменяться и численность особей в популяциях. Так, у насекомых и мелких растений открытых пространств численность особей в отдельных популяциях может достигать сотен тысяч и миллионов особей. Напротив, популяции животных и растений могут быть и сравнительно небольшими по численности. На одном из озер Помосковья численность популяции стрекоз Leucorrhinia albifrons достигала около 30 тыс. особей, численность популяций прыткой ящерицы Lacerta agilis — от нескольких сотен до нескольких тысяч особей, а численность популяций земляной улитки Cepea nemoralis — лишь 1000 экземпляров.
В связи с тем что любая популяция обладает строго определенной генетической, фенотипической, половозрастной и другой струк-турой,'она не может состоять из меньшего числа индивидов, чем необходимо для обеспечения стабильной реализации этой структуры и устойчивости популяции к факторам внешней среды (рис. 10.14).
В этом и состоит принцип минимального размера популяций. Минимальная численность популяций, обеспечивающая существование вида, является специфической для разных видов. Выход за пределы минимума грозит для популяции гибелью. Дальнейшее сокращение, например, тигра на Дальнем Востоке, неизбежно приведет к их автоматическому вымиранию из-за того, что оставшиеся единицы, не находя с достаточной частотой партнеров для размножения, вымрут на протяжении немногих поколений. В таком же положении могут оказаться и редкие растения, такие, как орхидея, «венерин башмачок» и другие.
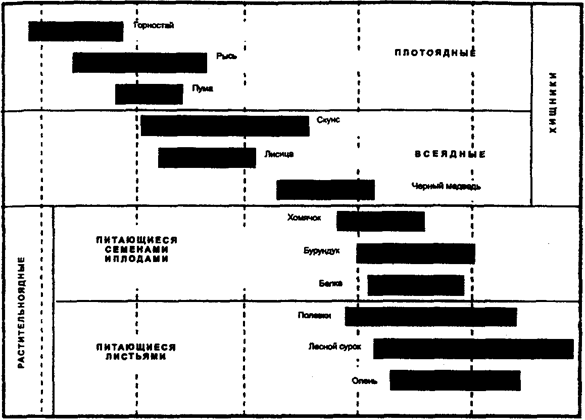
Рис. 10.14. Плотность популяций у разных видов
млекопитающих (в килограммах биомассы на гектар)
Закономерно предположить, что если есть минимум размера популяций, то возможен и максимум. Такое предположение помимо логической посылки основывается на соотношении законов максимума биогенной энергии и давления среды. Ю. Одум (1975) формирует закон как правило популяционного максимума. Популяции эволюционируют так, что регуляция их плотности осуществляется на значительно более низкой по сравнению с верхней асимптотой* емкости местообитания, достигаемой лишь в том случае, если полностью используются ресурсы энергии и пространства. При росте плотности популяции снижается обеспеченность пищей. У многих животных от потребления пищи прямо зависит плодовитость: при увеличении плотности популяции плодовитость падает, и это предотвращает дальнейший рост численности (рис. 10.15).
Правило популяционного максимума конкретизирует два обобщения. Первое из них известно как теория X. Г. Андреварты — Л. К. Бирча (1954), или теория лимитов популяциочной численности: численность естественных популяций ограничена истощением пищевых ресурсов и условий размножения, недоступностью этих ресурсов и слишком коротким периодом ускорения роста популяции. Второе обобщение дополняет первое и носит название теории биоценотической регуляции численности популяции К. Фридерикса (1927): регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида.
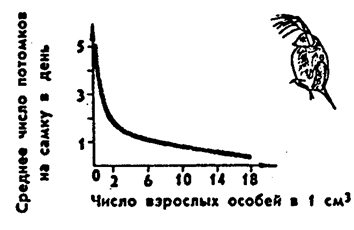
Рис. 10.15. Зависимость плодовитости у одного из видов
дафний (по Ю. Одуму, 1975)
Совокупность всех факторов, способствующих увеличению численности популяции, называется биотическим потенциалом. Несмотря на то что у разных видов составляющие биотического потенциала неодинаковы, имеется общее свойство: у всех видов он достаточно высок для стремительного увеличения численности при благоприятных условиях среды. Рост популяции может быть столь быстрым, что может привести к популяционному взрыву. Однако следует отметить, что повышение плотности популяций сверх оптимальной оказывает на них неблагоприятное воздействие, так как при этом иссякает кормовая база, сокращается жизненное пространство, появляются эпизоотии и т. д. (рис. 10.16).
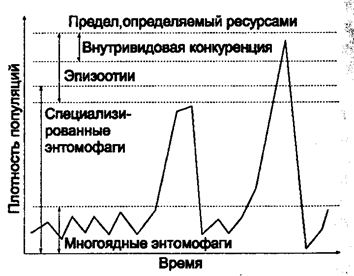
Рис. 10.16. Факторы, регулирующие численность
насекомых при различной плотности популяции
(по Г. А. Викторову, 1967)
Различают непериодические, редко наблюдаемые, и периодические, постоянные, колебания численности естественных популяций. К непериодическим колебаниям численности, а следовательно, и плотности популяции среди наземных организмов, могут быть отнесены вспышки массового размножения непарного шелкопряда Ocneria dispar в 1879 г. в южной и юго-восточной частях России, златогузки Euproctis chrysorrhoea, непарного и кольчатого шелкопрядов в период с 1948 по 1969 г. на Русской равнине и т. д. Резкий подъем численности нередко наблюдается у популяций, оказавшихся в новом местообитании. Примером этому может служить массовое размножение кроликов в Австралии, колорадского картофельного жука в Европе.
Периодические колебания численности популяций совершаются обычно в течение одного сезона или нескольких лет. Циклические изменения с подъемом численности в среднем через четыре года зарегистрированы у животных, обитающих в тундре, — леммингов, полярной совы, песца. Сезонные колебания численности характерны и для многих насекомых, мышевидных грызунов, птиц, мелких водных организмов.
В естественных условиях вероятность того, что все условия окажутся благоприятными для популяции, очень низка. Как правило, один или несколько абиотических (неоптимальная температура, кислотность, соленость, влажность) и биотических (присутствие хищников, паразитов, болезнетворных организмов, нехватка пищи) факторов становятся лимитирующими. Сочетание данных лимитирующих (ограничивающих) факторов называют сопротивлением среды (рис. 10.17).
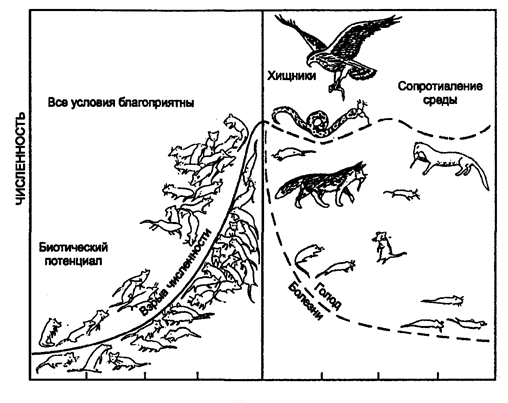
ПОКОЛЕНИЯ
Рис. 10.17. Сопротивление среды (гго Б. Небелу, 1993)
Сопротивление среды сильнее всего действует на молодых особей, больше других страдающих от нападения хищников, болезней, недостатка воды и пищи или других неблагоприятных условий.
Поддержание определенной численности или равновесное состояние получило название гомеостаза популяций. Рост, снижение или постоянство численности популяций зависит от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей).
Отсюда принцип изменения популяций можно сформулировать следующим образом: изменение популяции какого-либо вида — это результат нарушения равновесия между ее биотическим потенциалом и сопротивлением окружающей среды. Данное равновесие называют динамическим, или непрерывно регулирующимся, так как факторы сопротивления среды редко остаются неизменными в течение длительного времени. В какой-то год популяция может значительно снизить свою численность из-за засухи, а в последующие годы с нормальным увлажнением полностью восстановить ее. Подобные циклические колебания обычно продолжаются неопределенно долго. Это объясняет обобщающее правило максимума размера колебаний плотности популяционного населения. «Существуют определенные верхние и нижние пределы для средних размеров популяции, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени». Ю. Одум (1975) справедливо делает замечание, что может быть 100 птиц на 1 га и 20 000 почвенных членистоногих на 1 м2, но никогда не бывает 20 000 птиц на 1 м2 и 100 членистоногих на 1 га. Правило максимума размера колебаний плотности популяционного населения можно назвать законом количественной константности популяционного населения. Данная закономерность указывает на среднесистемное число особей популяции на единицу площади. Отклонение от этого закона, или правила, свидетельствует о неблагополучной ситуации в регионе, является биоиндикатором разлада в его экосистемах. Отсутствие или подавление численности какого-то вида в силу биотических и абиотических взаимосвязей определенно повлечет за собой цепь последствий, результаты которых следует предвидеть.
Дата добавления: 2016-02-24; просмотров: 1470;