Кора больших полушарий мозга
Новая кора (неокортекс) представляет собой слой серого вещества общей площадью 1500—2200 см2, покрывающий большие полушария; она составляет около 72 % всей площади коры и около 40 % массы головного мозга. В коре имеется около 14 млрд нейронов, количество глиальных клеток примерно в 10 раз больше. Кора большого мозга является в филогенетическом плане наиболее молодой нервной структурой; у человека она осуществляет высшую регуляцию функций организма и психофизиологические процессы, обеспечивающие различные формы поведения.
Нейронная организация новой коры.В направлении с поверхности в глубь коры различают 6 горизонтальных слоев.
I — молекулярный слой имеет очень мало клеток, но большое количество ветвящихся дендритов пирамидных клеток, формирующих сплетение, расположенное параллельно поверхности. На этих дендритах образуют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса.
II — наружный зернистый слой составлен в основном звездчатыми клетками и частично малыми пирамидными клетками. Волокна клеток второго слоя расположены преимущественно вдоль поверхности коры, образуя кортико-кортикальные связи.
III — наружный пирамидный слой состоит в основном из пирамидных клеток средней величины. Аксоны этих клеток, как и зернистые клетки II слоя, образуют кортикокортикальные ассоциативные связи.
IV внутренний зернистый слой по характеру клеток (звездчатые клетки) и расположению их волокон аналогичен наружному зернистому слою. В этом слое имеют синаптические окончания афферентные волокна, идущие от нейронов специфических ядер таламуса; здесь отмечена наибольшая плотность капилляризации.
V -- внутренний пирамидный слой образован средними и крупными пирамидными нейронами, причем в двигательной коре расположены гигантские пирамидные клетки Беца. Аксоны этих клеток образуют эфферентные кортико-спинальный и кортико-бульбарный двигательные тракты.
VI — слой полиморфных клеток образован преимущественно веретенообразными клетками, аксоны которых образуют кортико-таламические пути.
Оценивая в целом афферентные и эфферентные связи коры большого мозга, можно отметить, что в слоях I—IV происходит восприятие и обработка поступающих в кору сигналов. Напротив, покидающие кору эфферентные пути формируются преимущественно в V—VI слоях. Более детально деление коры на различные поля проведено на основе цитоархитектонических признаков (форма и расположение нейронов) К. Бродманом, который выделил 52 поля; многие из них характеризуются функциональными и нейрохимическими особенностями.
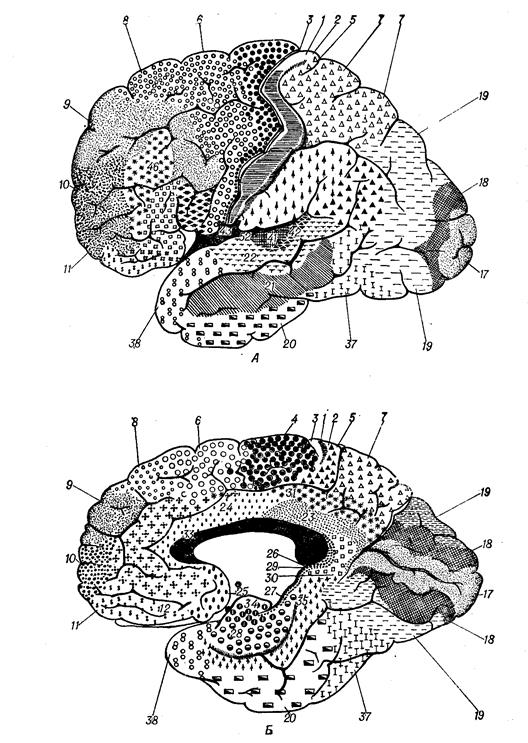
Цитоархитектоническая карта полей коры больших полушарий человека по К Бродману. А- латеральная, Б- медиальная поверхность.
Электрофизиологические исследования с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры показали, что все встречаемые на пути нейроны отвечают на раздражитель только одного качества, например, тактильный. Напротив, при погружении электрода под углом на его пути попадались нейроны разной модальности. Был сделан вывод, что в коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5—1,0 мм. Эти объединения были названы нейронными колонками. Они обнаружены в моторной коре, в различных зонах сенсорной коры. Соседние нейронные колонки могут взаимодействовать друг с другом.
Локализация функций в коре большогомозга интенсивно изучается в клинической и экспериментальной медицине, начиная с середины XIX в. При разработке этой проблемы были сформулированы две противоположные по смыслу концепции: узкого локализационизма и функциональной равноценности различных корковых структур. Современная концепция локализации функций базируется на принципе многофункциональности (но не равноценности) корковых полей.
Одним из наиболее известных вариантов функционального разделения коры большого мозга является выделение в ней сенсорной, ассоциативной и двигательной областей.
СЕНСОРНЫЕ ОБЛАСТИ КОРЫ
Это зоны, в которые проецируются сенсорные раздражители (проекционная кора, корковые отделы анализаторов). Они расположены преимущественно в теменной (поля 1—3), височной (поля 21, 22, 41, 42) и затылочной (поля 17—19) долях. Афферентные пути в сенсорную кору поступают преимущественно от релейных сенсорных ядер таламуса — вентральных задних латерального и медиального.
Зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и по-стоянные изменения чувствительности организма, называются первичными сенсорными областями (ядерные части анализаторов, по И.П. Павлову). Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, полимодальные нейроны которых отвечают на действие нескольких раздражителей.
Важнейшими сенсорными областями являются теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1—3). Эту зону обозначают как соматосенсорную область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата от мышечных, суставных, сухожильных рецепторов. Проекция головы и верхних отделов туловища находится в нижнелатеральных участках постцентральной извилины, проекция нижней половины туловища и ног - в верхнемедиальных зонах извилины, проекция нижней части голени и стоп расположена в коре парацентральной дольки на медиальной поверхности полушарий. При этом проекции наиболее чувствительных участков (язык, губы, гортань, пальцы) имеют относительно большие зоны по сравнению с другими частями тела. Предполагается, что в соматосенсорной области I в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности.
Кроме соматосенсорной области I, выделяют меньших размеров соматосенсорную область II, расположенную на границе пересечения центральной борозды с верхним краем височной доли, в глубине латеральной борозды. Степень локализации частей тела здесь выражена хуже. Функции области II плохо изучены. Известно, что сигналы в эту область поступают с обеих сторон тела и от других сенсорных областей мозга, например зрительных и слуховых.
Хорошо изученной первичной проекционной зоной является слуховая кора (поля 41, 42), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). В этой зоне в ответ на раздражение слуховых рецепторов кортиева органа формируются звуковые ощущения, различающиеся по громкости, тону и другим характеристикам. Здесь имеется четкая топическая проекция: в разных участках коры представлены различные участки кортиева органа. К проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и средней височных извилинах (поля 20 и 21). Обработанная сенсорная информация используется для формирования «схемы тела» и регуляции функций мозжечка (височно-мостомозжечковый путь).
Важнейшая первичная проекционная область новой коры расположена в затылочной коре – это первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки. Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Раздражение коры поля 17 приводит к возникновению световых ощущений. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные и слуховые раздражители.
Основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору.
АССОЦИАТИВНЫЕ ОБЛАСТИ КОРЫ
Ассоциативная (межсенсорная, межанализаторная кора) включает участки новой коры большого мозга, которые расположены рядом с сенсорными и двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций. Границы этих областей обозначены недостаточно четко, неопределенность преимущественно связана со вторичными (высшими) проекционными зонами, функциональные свойства которых являются переходными между свойствами первичных проекционных и ассоциативных зон. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и человека. У нас она составляет около 50 % всей коры и 70 % неокортекса.
Основной физиологической особенностью нейронов ассоциативной коры, отличающей их от нейронов первичных зон, является по-лисенсорность (полимодальность): они отвечают, как правило, не на один, а на несколько раздражителей. Полисенсорность нейронов ассоциативной коры создается кортико-кортикальными связями с разными проекционными зонами, зонами, связями с ассоциативными ядрами таламуса. В результате этого ассоциативная кора представляет собой своеобразный коллектор различных сенсорных возбуждений.
Таламолобная системапредставлена ассоциативными зонами лобной коры (поля 9— 14), имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса, других подкорковых ядер.
Основная роль лобной ассоциативной коры сводится к инициации базовых системных механизмов формирования функциональных систем целенаправленных поведенческих актов (П.К. Анохин).
В результате операции префронтальной лоботомии, при которой лобные доли по медицинским показаниям изолируются, наблюдаются выраженные изменения поведения, в котором обычно доминируют эмоциональные расстройства, неадекватность поступков и действий, особенно в изменившихся условиях.
Надо отметить, что некоторые ассоциативные центры включают в себя и участки височной коры (поле 39). В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины (поля 22, 37 и 42 левого доминантного полушария). Этот центр обеспечивает распознание и хранение устной речи, как собственной, так и чужой. В средней части верхней височной извилины (поле 22) находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей (поле 39) локализован центр чтения письменной речи, обеспечивающий распознание и хранение образов письменной речи.
Психические функции, осуществляемые ассоциативной корой, инициируют поведение организма, обязательным компонентом которого являются произвольные целенаправленные движения, осуществляемые при обязательном участии двигательной коры.
ДВИГАТЕЛЬНЫЕ ОБЛАСТИ КОРЫ
Представление о двигательной коре большого мозга начало формироваться с 80-х годов XIX в., когда было показано, что электрическое раздражение некоторых корковых зон у животных вызывает движение конечностей противоположной стороны. В двигательной коре выделяют первичную и вторичную моторные области. В первичной двигательной коре (прецентральная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела. При этом проекции мышц нижних конечностей и туловища расположены в верхних участках прецентральной извилины и занимают сравнительно небольшую площадь, а проекции мышц верхних конечностей, лица и языка расположены в нижних участках извилины и занимают большую площадь («двигательный человечек» Пенфилда). Двигательные реакции на раздражение первичной моторной коры осуществляются с минимальным порогом (высокая возбудимость): они представлены элементарными сокращениями мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральным). При поражениях этой корковой зоны утрачивается способность к тонким координированным движениям конечностей, особенно пальцев рук.
Вторичная двигательная к о р а (поле 6) расположена как на латеральной поверхности полушарий, впереди прецент-ральной извилины (премоторная кора), так и на медиальной поверхности, соответствующей коре верхней лобной извилины (дополнительная моторная область).
Вторичная двигательная кора в функциональном плане имеет главенствующее положение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1 с до начала движения. Кора поля 6 получает основную часть импульсации от базальных ганглиев и мозжечка, участвует в перекодировании информации о плане сложных движений. Раздражение коры поля 6 вызывает сложные координированные движения, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевые движения, а также музыкальный моторный центр (поле 45), тональности речи.
Эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные тракты, начинающиеся от гигантских пирамидных клеток Беца и менее крупных пирамидных клеток V слоя коры прецентральной извилины (60 % волокон), премоторной коры (20 % волокон) и постцентральной извилины (20 % волокон). От крупных пирамидных нейронов формируются преимущественно пирамидные пути. Они иннервируют крупные (фазические, высокопороговые) α-мотонейроны в моторных центрах ствола и спинного мозга, регулирующие движение тела и его частей в пространстве. Крупные пирамидные клетки имеют фоновую импульсную активность около 5 имп/с, которая при движениях увеличивается до 20—30 имп/с. От мелких пирамидных клеток формируются преимущественно экстрапирамидные пути. Эти клетки имеют фоновую активность около 15 имп/с, которая во время движения увеличивается или уменьшается; они иннервируют мелкие (тонические, низкопороговые) α-мотонейроны в стволовых и спинальных двигательных центрах и регулируют тонус мышц и позу.
Волокна пирамидного пути оканчиваются на α-мотонейронах двигательных ядер III— VII и IX—XII черепных нервов (кортико-бульбарный тракт) или спинальных двигательных центрах (кортико-спинальные тракты). Через двигательную кору и пирамидные пути осуществляются произвольные простые движения и сложные целенаправленные двигательные программы (например, профессиональные навыки), формирование которых начинается в базальных ганглиях и мозжечке и заканчивается во вторичной моторной коре. Большинство волокон пирамидных путей осуществляет перекрест, однако небольшая часть волокон остается неперекрещенными, что способствует компенсации нарушения функции движения при односторонних поражениях. Через пирамидные тракты осуществляет свои функции и премоторная кора. К ним относятся двигательные навыки письма, сочетанные повороты головы, глаз и туловища, речевые функции (речедвигательный центр Брока, поле 44). В регуляции письменной и особенно устной речи имеется выраженная асимметрия больших полушарий мозга: у 95 % правшей и у 70 % левшей устная речь контролируется левым полушарием.
К корковым экстрапирамидным путям относятся кортико-рубральные и кортико-ре-тикулярные тракты, начинающиеся приблизительно от зон, которые дают начало пирамидным путям. Волокна кортико-рубрального тракта оканчиваются на нейронах красных ядер среднего мозга, от которых далее идут руброспинальные тракты. Волокна кортико-ретикулярных трактов оканчиваются на нейронах медиальных ядер ретикулярной формации моста (от них идут медиальные ретикулоспинальные тракты) и на нейронах ретикулярных гигантоклеточных ядер продолговатого мозга, от которых начинаются латеральные ретикулоспинальные тракты. Через эти тракты осуществляется регуляция тонуса и позы, обеспечивающих точные, целенаправленные движения. Корковые экстрапирамидные тракты являются компонентом экстрапирамидной системы головного мозга, к которой относятся мозжечок, базальные ганглии, моторные центры ствола; она осуществляет регуляцию тонуса, позы, координацию и коррекцию движений. Поскольку кортико-пирамидные пути отдают многочисленные коллатерали к структурам экстрапирамидной системы, обе системы работают в функциональном единстве, и некоторые авторы [Шмидт Р., 1985] считают нецелесообразным их разделение. О функциональном единстве этих систем свидетельствуют и опыты по перерезке пирамидных путей у приматов. У таких животных не было отмечено грубых нарушений движений: сохранялись стояние, ходьба, двигательные условные рефлексы, сложные программированные движения. Не восстанавливались только «специфические» функции пирамидных путей — независимые, точные движения пальцев рук и контроль силы этих движений.
Определяющую роль в структурном обеспечении двигательных актов играют следующие образования:
• фронтальная кора, выступающая в качестве инициатора замысла действия;
• ассоциативная теменная кора, выполняющая функции интеграции гетеромодальных возбуждений, а также командные функции в отношении ряда параметров движения;
• базальные ганглии, мозжечок, возможно, выполняющие функции накопителей «двигательных программ»;
• моторная кора, моторные центры ствола, спинного мозга, обеспечивающие выбор мышечных эффекторов в ходе реализации программы двигательного акта и управления двигательным процессом на базе обратной афферентации.
У человека четко проявляются межполушарные взаимотношения в двух главных формах — функциональной асимметрии больших полушарий и совместной их деятельности.
Функциональная асимметрия полушарийявляется важнейшим психофизиологическим свойством головного мозга человека. Ее исследование началось в середине прошлого века, когда французские медики М. Дакс (1836) и П. Брока (1861) показали, что нарушение речи человека возникает при поражении коры нижней лобной извилины, как правило, левого полушария. Несколько позже немецкий психиатр К. Вернике (1874) обнаружил в коре заднего отдела верхней височной извилины левого полушария слуховой центр речи, поражение которого приводит к нарушению понимания устной речи. Эти результаты и наличие моторной асимметрии (праворукость) сформировали концепцию, согласно которой для человека характерно левополушарное доминирование, образовавшееся эволюционно в результате трудовой деятельности и являющееся специфическим свойством его мозга. Однако в XX в. в результате применения различных методических подходов, особенно при исследовании больных с расщепленным мозгом (перерезка мозолистого тела), было показано, что по ряду психофизиологических функций доминирует не левое, а правое полушарие, и возникла концепция частичного доминирования полушарий.
Выделяют психическую, сенсорную и моторную межполушарные функциональные асимметрии мозга.
Парность в деятельности больших полушарий обеспечивается наличием комиссуральной системы — мозолистого тела, передней, задней, гиппокампальной и хабенулярной комиссур, межбугрового сращения, которые анатомически соединяют два полушария головного мозга. Результаты, свидетельствующие о переносе возбуждения из одного полушария в другое, получены преимущественно с использованием электрофизиологических методик и метода условных рефлексов.
Основные факты, полученные с помощью электрофизиологических методик, показали, что возбуждение из участка раздражения одного полушария передается через комиссуральную систему не только в симметричный участок другого полушария, но также и в несимметричные участки коры (например, из зрительной коры одного полушария в моторную кору противоположного полушария).
ЭЛЕКТРОЭНЦЕФАЛОГРАФИЧЕСКИЙ МЕТОД
Электроэнцефалографический метод основан на регистрации суммарной электрической активности мозга – электроэнцефалограммы (ЭЭГ). Впервые ЭЭГ у животных была зарегистрирована В.В. Правдич-Неминским (1913), у человека -- Г. Бергером (1929). Запись ЭЭГ возможна как с поверхности кожи головы, так и непосредственно с коры головного мозга. В последнем случае она называется электрокортикограммой (ЭКоГ).
Регистрация ЭЭГ производится с помощью биполярных (оба активны) или униполярных (активный и индифферентный) электродов, накладываемых на проекции лобных, центральных, теменных, височных и затылочных областей головного мозга. В клинике обычно используется запись с помощью 10—12 электродов. Основными анализируемыми параметрами ЭЭГ являются частота и амплитуда волновой активности. Кроме записи фоновой ЭЭГ, у испытуемых часто регистрируют изменения ЭЭГ при действии тех или иных раздражителей экстероцептивных (световых, звуковых и др.), проприоцептивных, вестибулярных и др.
Происхождение волн ЭЭГ выяснено недостаточно. Наиболее вероятно, что ЭЭГ отражает алгебраическую сумму возбуждающих и тормозных постсинаптических потенциалов (ВПСП и ТПСП) множества нейронов в зоне расположения отводящих электродов. На ЭЭГ регистрируется 4 основных физиологических ритма: альфа, бета, тета и дельта.
Альфа-ритмимеет частоту 8—13 Гц, амплитуду до 70 мкВ. Этот ритм наблюдается у человека в состоянии физического, интеллектуального и эмоционального покоя. Альфа-ритм является упорядоченным регулярным ритмом. Если он доминирует, ЭЭГ оценивается как синхронизированная. Механизм синхронизации ЭЭГ связан с деятельностью выходных ядер таламуса.
Альфа-ритм преобладает у 85—95 % здоровых людей старше девятилетнего возраста. Лучше всего он выражен в затылочных областях мозга, в передних (лобной и центральной) областях, часто сочетается с бета–ритмом. Вариантом альфа-ритма являются «веретена сна» длительностью 2—8 с, которые наблюдаются при засыпании и представляют собой регулярные чередования нарастания и снижения амплитуды волн в частотах альфа-ритма.
Бета-ритмимеет частоту 14—30 Гц, амплитуду до 30 мкВ, характеризуется нерегулярными по частоте низкоамплитудными волнами, которые сменяют альфа-ритм при сенсорной стимуляции (например, при действии света, сильного звука), при эмоциональном возбуждении. Наиболее выражен бета-ритм в лобных, центральных областях головного мозга. Смена альфа-ритма бета-ритмом называется десинхронизацией ЭЭГ. Ее механизм связывают с активирующим влиянием на кору большого мозга восходящей ретикулярной формации ствола и лимбической системы. Бета-ритм отражает высокий уровень функциональной активности головного мозга.
Тета-ритмимеет частоту 4—7 Гц, амплитуду — до 200 мкВ. У бодрствующего человека тета-ритм на ЭЭГ регистрируется обычно в передних областях мозга при длительном эмоциональном напряжении. Отчетливо проявляется у детей, пребывающих в состоянии эмоции неудовольствия. Тета-ритм почти всегда выявляется в процессе развития фаз медленноволнового сна.
Дельта-ритмимеет частоту 0,5—3,0 Гц, амплитуду — 200—300 мкВ. Эпизодически регистрируется во всех областях головного мозга. Стабильно фиксируется во время глубокого медленноволнового сна. Появление этого ритма у бодрствующего человека свидетельствует о снижении функциональной активности мозга.
Происхождение тета- и дельта-ритмов ЭЭГ связывают с активностью соответственно мостовой и бульбарной синхронизирующих систем ствола мозга.
Дата добавления: 2016-02-20; просмотров: 2178;