МЕХАНИЗМЫ СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ МЫШЕЧНОГО ВОЛОКНА
При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0.05 с (50 мс). За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0.5 мс. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный им пульс вызывает перемещение синаптических пузырьков к пресинаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается ацетилхолинэстеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако, при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервно-мышечный синапс. Эти процессы лежат в основе периферических механизмов утомления при длительной и тяжелой мышечной работе.
Выделившийся всинаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Небольшое подпороговое раздражение вызывает лишь местное возбуждение небольшой амплитуды — потенциал концевой пластинки (ПКП).
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью 5 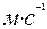 ) распространяется вдоль по поверхности мышечного волокна и заходите поперечные
) распространяется вдоль по поверхности мышечного волокна и заходите поперечные
трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматического ретикулума ионов Са 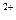 , которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
, которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
Под влиянием Са длинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е. механическую реакцию мышечного волокна (рис. 10).
Энергия гребкового движения одного мостика производит перемещение на 1 % длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са . Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФ-азы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению
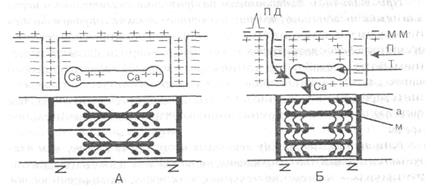
Рис. 10. Схема электромеханической связи в мышечном волокне
На А: состояние покоя, на Б — возбуждение и сокращение
да — потенциал действия, мм — мембрана мышечного волокна,
п _ поперечные трубочки, т — продольные трубочки и цистерны с ионами
Са ,а — тонкие нити актина, м — толстые нити миозина
с утолщениями (головками) на концах. Зет-мембранами ограничены
саркомеры миофибрилл. Толстые стрелки — распространение потенциала
действия при возбуждении волокна и перемещение ионов Са из цистерн
и продольных трубочек в миофибриллы, где они содействуют образованию
мостиков между нитями актином и миозином и скольжение этих нитей
(сокращение волокна) за счет гребковых движений головок миозина.
имеющихся мостиков и образованию в присутствии Са новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна — через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма — «кальциевого насоса», который обеспечивает откачку ионов Са из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
Дата добавления: 2016-02-10; просмотров: 859;