Строение клеток бактерий
Большинство бактерий является одноклеточными организмами. Эволюция выработала у них такие адаптивные механизмы, которые позволяют каждой отдельной клетке осуществлять все необходимые жизненные функции, рост, развитие и размножение.
Бактериальная клетка содержит до 80 % воды. Остальная часть представлена белками, полисахаридами, липидами, нуклеиновыми кислотами. В сухом веществе клетки они соответственно составляют в среднем 52, 18 и19 %.
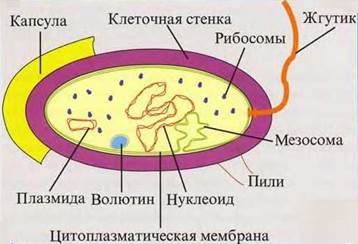
Цитоплазму, окруженную мембраной, называют протопластом. Снаружи от ЦПМ располагаются поверхностные структуры прокариотной клетки: клеточная стенка, слизистые капсулы или чехлы, жгутики, ворсинки.
 В связи с особенностями строения клеточной стенки бактерии делят на грамположительные и грамотрицательные. Название происходит от способа окрашивания, предложенного датским ученым Х. Грамом: фиксированные клетки бактерий обрабатывают сначала реактивом кристаллическим фиолетовым, затем йодом, вследствие чего появляется специфическое окрашивание. При последующей обработке этиловым спиртом у одних бактерий окрашивание исчезает – это грамотрицательные бактерии. Другие бактерии сохраняют окрашенный комплекс, поэтому получили название грамположительных бактерий.
В связи с особенностями строения клеточной стенки бактерии делят на грамположительные и грамотрицательные. Название происходит от способа окрашивания, предложенного датским ученым Х. Грамом: фиксированные клетки бактерий обрабатывают сначала реактивом кристаллическим фиолетовым, затем йодом, вследствие чего появляется специфическое окрашивание. При последующей обработке этиловым спиртом у одних бактерий окрашивание исчезает – это грамотрицательные бактерии. Другие бактерии сохраняют окрашенный комплекс, поэтому получили название грамположительных бактерий.
Клеточная стенка прокариот устроена очень сложно. Основная масса клеточной стенки грамположительных бактерий состоит из гетерополимерного соединения – пептидогликана. Наряду с пептидогликаном в клеточной стенке грамположительных бактерий содержатся полимерные соединения – тейхоевые кислоты, в молекулы которых входит спирт рибит или глицерин. Кроме того, в клеточной стенке этих бактерий в небольших количествах представлены полисахариды, белки и липиды. Все три класса соединений, образующих клеточную стенку, связаны между собой ковалентными связями и представляют собой очень прочную упорядоченную структуру.
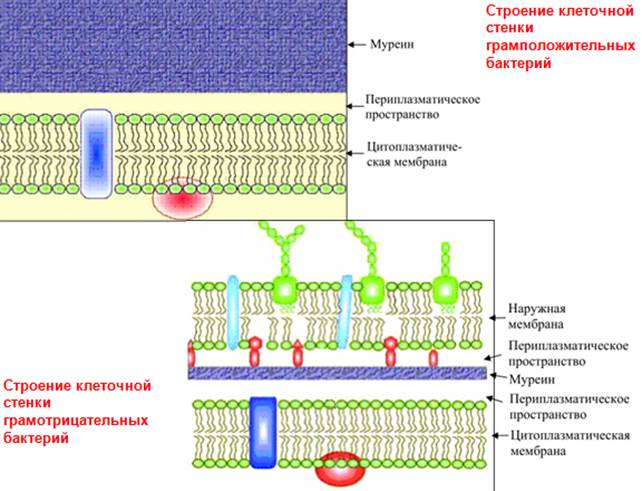
Клеточная стенка у грамотрицательных бактерий устроена еще более сложно. Она состоит из нескольких слоев: внутренний тонкий слой образован пептидогликаном, наружный слой представляет собой мембрану, состоящую из белков, липидов и полисахаридов. Между ними, в периплазматическом пространстве, расположен рыхлый слой белков.
Некоторые бактерии имеют отличия в строении клеточной стенки (метанообразующие, галофильные), другие не имеют ее вовсе (микоплазмы).
Клеточная стенка выполняет ряд физиологических функций: механическую; защитную; транспортную; служит местом локализации ряда веществ, например ферментов у грамотрицательных бактерий в периплазматическом пространстве между двумя мембранами и др.
Защитной функцией обладают и слизистые образования поверхностных структур, состоящие главным образом из особых полисахаридов и реже из полипептидов. Они же могут служить местом запаса воды и питательных веществ и одним из способов взаимной связи между соседними клетками у колониальных микроорганизмов.
Многие прокариоты способны к активному движению. Направленные движения микроорганизмов называют таксисами. В зависимости от действующего фактора различают хемо-, аэро-, фототаксисы и др. У разных микробов наблюдаются различные формы движения:
1. Жгутиковое. Жгутики представляют собой многократно изогнутые длинные структуры, которые прикреплены к ЦПМ. В состав жгутиков входит специфический белок флагеллин. Энергия для движения с помощью жгутиков получается при превращении градиента электрохимического потенциала в механическую работу.
2. Извивание. Например, у спирохет вокруг протопласта обвивается аксиальная (опорная) нить, состоящая из тех же веществ, что и жгутики. Движение спирохет происходит за счет вращения аксиальных фибрилл в периплазматическом пространстве между пептидогликановым слоем и наружной мембраной. Это вращение создает на поверхности клеточной стенки эластичную волну, благодаря которой спирохеты могут быстро вращаться вокруг своей длинной оси, извиваться и передвигаться в жидкой среде по винтовому или волнообразному способу.
3. Скольжение. Некоторые бактерии, не обладающие жгутиками, передвигаются по твердому субстрату за счет скольжения (миксобактерии, нитчатые серобактерии и др.). Это слизеобразующие микроорганизмы. Объяснение механизма движения этих бактерий видят в гипотезе бегущей волны.
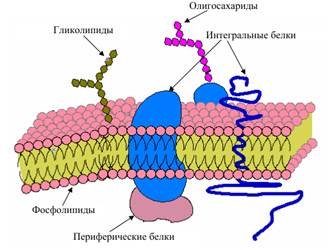 Цитоплазматическая мембрана (ЦПМ) у многих бактерий является единственной мембраной. На ее долю приходится 8 – 15 % сухой массы клетки. У фототрофных микроорганизмов, использующих энергию солнечного света, и хемоавтотрофных, пользующихся энергией окисления химических веществ, формируются мембранные структуры, тесно связанные с ЦПМ и выполняющие функции фотосинтеза и хемосинтеза. Они получили название внутрицитоплазматических мембран. Химический состав ЦПМ у грамположительных и грамотрицательных бактерий и эукариот практически одинаков.
Цитоплазматическая мембрана (ЦПМ) у многих бактерий является единственной мембраной. На ее долю приходится 8 – 15 % сухой массы клетки. У фототрофных микроорганизмов, использующих энергию солнечного света, и хемоавтотрофных, пользующихся энергией окисления химических веществ, формируются мембранные структуры, тесно связанные с ЦПМ и выполняющие функции фотосинтеза и хемосинтеза. Они получили название внутрицитоплазматических мембран. Химический состав ЦПМ у грамположительных и грамотрицательных бактерий и эукариот практически одинаков.
Цитоплазматическая мембрана бактерий выполняет много функций.
1. барьерную;
2. здесь находятся ферменты, с участием которых происходит образование различных компонентов клеточной стенки;
3. в ЦПМ расположены электронтранспортные цепи фотосинтеза, хемосинтеза, дыхания, анаэробного дыхания и ферментные комплексы, обеспечивающие превращение электрохимической энергии в энергию макроэргов АТФ;
4. ЦПМ участвует в процессах передачи наследственных свойств благодаря закреплению на ней кольцевой ДНК клетки, в поглощении воды и элементов питания.
Типичных органелл, т.е. структур, обособленных от цитоплазмы, у прокариот нет. Среди внутрицитоплазматических мембран бактерий следует отметить мезосомы, выполняющие функции энергообмена, а также выделения (секреторные функции). Предположительно с мезосомами связывают компартментацию клеток, обеспечивающую определенную последовательность в ходе различных ферментативных реакций, доставку субстратов для синтеза клеточной стенки. К внутрицитоплазматическим структурам относятся тилакоиды пурпурных бактерий, хлоросомы зеленых бактерий и фикобилисомы цианобактерий, выполняющие фотосинтетическую функцию.
Хлоросомы в виде продолговатых пузырьков длиной 90–150 нм и шириной 25–75 нм окружены однослойной мембраной толщиной 2 – 3 нм, состоящей только из молекул белков. Хлоросомы плотно примыкают к ЦПМ. В них содержатся бактериохлорофиллы, роль которых заключается в улавливании квантов света.
Фикобилисомы имеют вид гранул диаметром 25 – 55 нм. В них располагаются пигменты белковой природы фикобилипротеиды, растворимые в воде. Они также способны улавливать энергию солнечного света.
Цитоплазма, состоящая из белков, продуктов обмена веществ и различных субстратов, носит название цитозоля. Другая часть, структурная, содержит генетический аппарат, рибосомы, а у некоторых микроорганизмов и внутрицитоплазматические мембраны, и различные включения. Вся цитоплазма насыщена рибосомами, где осуществляются, как и у высших растений, синтез белка. Рибосомы отличаются от рибосом эукариот своими меньшими размерами. Число рибосом в бактериальных клетках зависит от интенсивности биосинтеза белка в них.
Генетический аппарат прокариотной клетки представлен, как правило, одной молекулой ДНК, имеющей форму кольца и названной бактериальной хромосомой. Впечатляют ее размеры: в развернутом виде ее длина почти в 1000 раз превышает длину прокариотной клетки. Бактериальная хромосома – высокоупорядоченная, компактная и суперспирализованная структура, имеющая у большинства бактерий молекулярную массу в пределах 1 – 3 · 109.
Молекула ДНК располагается в определенном месте цитоплазмы и не отделена от нее мембраной, как у эукариот. Поэтому, в отличие от последних, генетический аппарат прокариот называют нуклеоидом, а не ядром. Гистоны у большинства бактерий отсутствуют. Лишь недавно гистоноподобные белки, связанные с ДНК, обнаружены у некоторых микоплазм и некоторых цианобактерий. У каждого вида содержание пар азотистых оснований А + Т и Г + Ц в молекуле ДНК постоянно и служит важным диагностическим признаком.
Удвоение молекул ДНК предшествует делению прокариотной клетки, и начинается в точке прикрепления кольцевой хромосомы к ЦПМ. Здесь расположены ферменты, которые участвуют в процессах репликации ДНК. Водородные связи между полинуклеотидными нитями разрываются, и на однонитиевых участках, как на матрице, синтезируются комплементарные цепи дочерних молекул ДНК. Параллельно с репликацией ДНК происходит рост мембраны на участке контакта ДНК с ЦПМ, приводя к разделению вновь образовавшихся молекул ДНК и оформлению их в самостоятельные хромосомы двух дочерних бактериальных клеток.
Дата добавления: 2016-01-30; просмотров: 2301;