Добавочный и подъязычный нервы
Добавочный и подъязычный нервы чисто двигательные нервы. Добавочный нерв имеет веточки, выходящие из шейных сегментов спинного мозга.
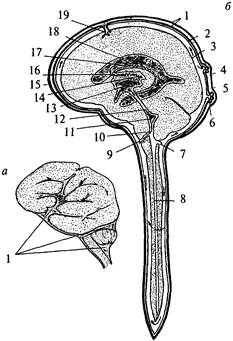
Структурная организация продолговатого мозга отличается от организации спинного мозга. Скопления клеток – ядра продолговатого мозга – перемежаются с пучками волокон, которые не всегда имеют поверхностное расположение. Чувствительные (сенсорные) ядра черепно-мозговых нервов располагаются дорсолатерально, двигательные (моторные) – вентрально, вегетативные ядра занимают промежуточное положение. Кроме ядер черепно-мозговых нервов в продолговатом мозге расположены ядра, на которых переключается сенсорная информация (ядра олив, ядра Голля и Бурдаха), и ядра ретикулярной формации, а также дыхательный, сосудодвигательный и пищеварительные центры (рис. 28).
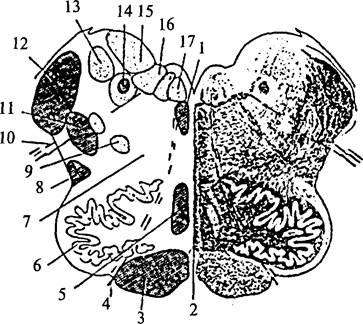
| Рис. 28. Поперечный разрез продолговатого мозга Левая половина – схема расположения ядер (обозначены светлым) и проводящих путей (обозначены темным): 1 – задний продольный пучок; 2 – шов; 3 – пирамида; 4 – корешок подъязычного нерва; 5 – медиальная петля (осязательный путь); 6 – ядро оливы; 7 – ретикулярная формация; 8 – спиноталамический тракт; 9 – двойное ядро; 10– корешок блуждающего нерва; 11 – дорсальный продольный пучок; 12 – нижние ножки мозжечка; 13 – ядро Дейтерса; 14 – одиночный (солитарный) тракт и его ядро; 15 – верхнее вестибулярное ядро; 16 – дорсальное ядро блуждающего нерва; 17 – ядро подъязычного нерва |
В продолговатом мозге и других стволовых отделах мозга (Варолиевом мосту и среднем мозге) расположена особая клеточная структура – ретикулярная формация (рис. 28, 7). В функциональном отношении ретикулярная формация ствола представляет собой единую структуру. Название этой структуры отражает ранние представления о том, что отдельные нейроны ретикулярной формации имеют обширные связи друг с другом и образуют нечто подобное нейропилю, в котором возбуждение распространяется диффузно, подобно тому, как это происходит в нервной системе кишечнополостных. Однако в дальнейшем эти представления не подтвердились.
Четких границ между отдельными ретикулярными и неретикулярными группами клеток не установлено, однако выделено до 98 ядерных групп, относящихся к ретикулярной формации. Основными ядрами являются: ядра шва и гигантоклеточное ядро продолговатого мозга, центральное и ретикулярное ядра моста.
Клетки ретикулярной формации очень разнообразны по форме и величине. Для них характерно наличие значительно разветвленного дендритного дерева и длинных аксонов. Афферентные входы ретикулярная формация получает как по коллатералям восходящих (сенсорных) путей, так и от вышележащих структур, в том числе от коры больших полушарий и мозжечка. Таким образом, ретикулярная формация интегрирует влияние большого числа мозговых структур. В свою очередь сама она оказывает влияние как на вышележащие, так и на нижележащие структуры.
Нисходящие и восходящие волокна ретикулярной формации покидают ядра на всем ее протяжении, не имея четких пространственных разграничений. Кроме того, существуют аксоны, которые Т-образно ветвятся. Одна из ветвей идет вниз, а другая – в верхние отделы головного мозга.
Нисходящие волокна образуют ретикулоспинальный тракт. Через него ретикулярная формация оказывает влияние как на двигательную деятельность спинного мозга (осуществление спинальных рефлексов), так и на вегетативную регуляцию (сосудодвигательная, дыхательная, пищеварительная функции). Ретикулярная формация воздействует на соматические и вегетативные центры в двух противоположных направлениях: торможения и возбуждения.
Восходящие влияния ретикулярной формации направлены на регуляцию деятельности коры больших полушарий. Большинство ретикулярных волокон достигает коры через переключение в неспецифических ядрах таламуса. Действие восходящих ретикулярных влияний заключается в широкой активации корковых структур. Торможение активности ретикулярной формации ведет к наступлению сна, активация ее приводит к реакции пробуждения.
Характерной особенностью нейронов ретикулярной формации является их высокая чувствительность к химическим факторам. Такие факторы, как уровень углекислого газа и кислорода, содержание адреналина, ацетилхолина и серотонина, относительно небольшие концентрации фармакологических веществ, изменяют активность нейронов ретикулярной формации, а вместе с этим и ее влияние на кору больших полушарий, соматические и вегетативные рефлексы.
Филогенез.
Филогенез.Вфилогенезе развитие продолговатого мозга определяется развитием чувствительных органов боковой линии, слуха, тактильной чувствительности тела. У всех позвоночных животных, кроме млекопитающих, в продолговатом мозге расположены ядра V– XII пар нервов. Развитие органов боковой линии и слуха определяет утолщение дорсальной части боковой стенки продолговатого мозга. У человека из этих структур остаются слуховые бугорки.
Вентральные структуры продолговатого мозга развиваются благодаря связям вышележащих отделов (коры больших полушарий) с двигательными структурами спинного мозга. Поэтому у низших позвоночных пирамиды отсутствуют, а у млекопитающих становятся ясно различимы. Оливы появляются только у высших обезьян. Поскольку наибольший кортикальный контроль над двигательными структурами наблюдается у человека, у него же наиболее выражены и пирамиды, и оливы.
Филогенетически ретикулярная формация является древней структурой. У низших позвоночных она распространена по всей центральной нервной системе. У высших позвоночных эта структура хорошо представлена только в стволовых отделах мозга, так как функцию регуляции берут на себя конечный мозг и мозжечок.
Онтогенез.
Онтогенез.Вонтогенезе продолговатый мозг является производным ромбовидного мозгового пузыря (rhombencephalon), который делится на задний (metencephalon) и добавочный мозг (myelencephalon). Полость ромбовидного мозга не делится и остается общей для продолговатого и заднего мозга – ГУ желудочком. Добавочный мозг развивается в продолговатый. Продолговатый мозг формируется так же, как и спинной. С дорсальной стороны он имеет крыловидные пластинки, между ними крышу, с вентральной стороны – базальные пластины и дно. Однако крыловидные пластинки расходятся, крыша растягивается и становится очень тонкой. В ней образуется сосудистое сплетение, которое заметно уже на 6-й неделе развития.
В структурах продолговатого мозга формируются ядра черепно-мозговых нервов. Причем почти все ядра нервов (тройничного, отводящего, лицевого, слухового), принадлежащие во взрослом состоянии мосту, закладываются первоначально в ростральных отделах продолговатого мозга, и только позже они перемещаются в структуры заднего мозга.
Сосудистое сплетение IV желудочка на начальных этапах эмбрионального развития человека представляет собой сплошную пластинку. В дальнейшем она прорывается, образуя отверстия: места сообщения между полостью IV желудочка и полостью подпаутинного пространства.
Задний мозг
Задний мозг развивается из заднемозгового пузыря (те tencéphalon), являющегося производным ромбовидного пузыря (rhombencephalon). Вентральная часть заднего мозга представляет собой продолжение стволовых структур и называется Варолиев мост. Варолиев мост несет в своем составе полость заднего мозга – часть ромбовидной ямки. Крыша ромбовидной ямки претерпевает значительные изменения и развивается в мозжечок, представляющий собой дорсальный вырост заднего мозга.
Варолиев мост
Варолиев мост (pons Varolii) – это вентральная часть заднего мозга. Сам мост образует структуры ростральной части дна IV желудочка. Дорсальная поверхность моста представляет собой верхний треугольник ромбовидной ямки. Полость ромбовидной ямки рострально сужается и переходит в водопровод среднего мозга. Сверху полость ромбовидной ямки прикрыта верхним мозговым парусом, который вместе с нижним мозговым парусом и сосудистым сплетением образует крышу IV желудочка, имеющую вид шатра. Латеральные стенки IV желудочка в области моста образованы средними и верхними ножками мозжечка (см. рис. 25, 33).
Вентральная поверхность Варолиева моста представляет собой мощную поперечно-волокнистую выпуклость белого вещества. По центру вентральной поверхности моста проходит глубокая борозда – канавка основной артерии мозга (suclus basil laris). Латерально вентральная выпуклость переходит в мощные средние ножки мозжечка (см. рис. 25, 5; 27, 4).
От Варолиевого моста отходят четыре пары черепно-мозговых нервов (см. рис. 26; 27).
V – тройничный нерв (п. trigeminus);
VI – отводящий нерв (п. abducens);
VII – лицевой нерв (п. facialis);
VIII – предверно-улитковый, или слуховой, нерв (п. vestibulo cochlearis).
На поперечных срезах, как и в продолговатом мозге, видны белое вещество и ядра серого вещества. Поперечные волокна, составляющие трапециевидное тело, делят толщу моста на более крупную вентральную (основание моста) и дорсальную (покрышка моста) части. В вентральной части преобладает белое вещество проводящих путей, являющихся продолжением проводящих путей ножек среднего мозга. Серое вещество вентральной части моста образует собственные ядра моста (ядра основания моста). В этих ядрах заканчиваются идущие из коры больших полушарий нисходящие кортикомостовые пути и коллатерали от кортикоспинальных путей. От собственных ядер моста отходят волокна, которые переходят на противоположную сторону и образуют трапецевидные тела, переходящие в средние ножки мозжечка.
Дорсальная часть моста является непосредственным продолжением продолговатого мозга. В ней располагаются переключательные ядра сенсорных систем, ядра черепно-мозговых нервов и ретикулярной формации.
В филогенезе
В филогенезеу низших позвоночных животных Варолиев мост не отделен четко от продолговатого мозга. Он обособляется только у млекопитающих. Это происходит с развитием коры и нисходящих из нее проекционных путей. Вместе с этим растет количество собственных ядер вентральной части моста. Это обусловливает появление и развитие средних ножек мозжечка и его полушарий. Особенно значительно вентральные части моста и средние ножки мозжечка выражены у человека.
В онтогенезе
В онтогенеземост, как отдел заднего мозга, происходит из ромбовидного мозгового пузыря. На стадии пяти мозговых пузырей ромбовидный мозг делится на добавочный (myelencephalon), из которого развивается продолговатый мозг, и задний мозг (те tencéphalon). Крыша заднего мозга преобразуется в мозжечок, а дно и стенки его становятся структурами моста. Полость ромбовидного мозга остается общей для продолговатого мозга и моста и является полостью IV желудочка.
Почти все ядра черепно-мозговых нервов моста закладываются в ростральных отделах продолговатого мозга. Их перемещение в мост происходит после образования мозгового изгиба. На 7-й неделе развития эмбриона клетки крыловидной пластинки продолговатого мозга мигрируют в ростровентральном направлении и образуют на вентральной поверхности моста понтобульбарное тело, которое позже превращается в собственные ядра моста.
Мозжечок
Мозжечок (cerebellum) располагается на дорсальной поверхности ствола мозга. Вентральная поверхность мозжечка прилежит к парусам IV желудочка и тесно связана со стволовыми структурами тремя парами мозжечковых ножек: с продолговатым мозгом нижними ножками (pedunculus cerebellaris inferior) (рис. 29, 7), с мостом средними ножками (pedunculus cerebellaris médius) (рис. 29, 2) и со средним мозгом – верхними ножками (pedunculus cerebellaris superior) (рис. 29, 1). Наиболее мощными являются средние ножки. Все ножки выходят из мозжечка рядом, а потом верхние ножки направляются к среднему мозгу вместе с верхним мозговым парусом, а нижние – к продолговатому мозгу вместе с нижним мозговым парусом. Рострально над мозжечком лежат затылочные доли большого мозга, которые заходят за его дорсальные границы мозжечка. Мозжечок отделен от большого мозга глубокой поперечной щелью мозга. Он, как и большой мозг, покрыт тремя оболочками.
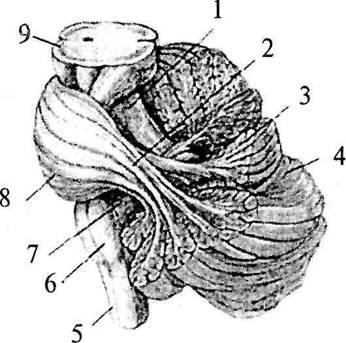
| Рис. 29. Расположение мозжечка на стволе мозга (часть ткани мозжечка убрана): 1 – верхняя ножка мозжечка; 2 – средняя ножка мозжечка; 3 – лист (долька мозжечка); 4 – левое полушарие мозжечка; 5 – спинной мозг; 6 – продолговатый мозг; 7 – нижняя ножка мозжечка; 8 – мост; 9 – средний мозг |
Анатомически мозжечок человека состоит из трех основных частей: двух полушарий (hemi - spheria cerebelli) и соединяющей их средней части – червя (vermis cerebélli). Поверхность мозжечка изрезана глубокими ветвящимися бороздами. Глубокие борозды мозжечка делят полушария и червь на дольки, которые объединяются в доли: верхние, задние и нижние. Доли отделяются друг от друга щелями.
Между обоими полушариями мозжечка располагается изрезанная узкими, параллельно идущими бороздами средняя часть – червь (см. рис. 30а, 13). На нем различают верхнюю поверхность – верхний червячок, и нижнюю – нижний червячок. Две продольно идущие бороздки на каждой поверхности мозжечка отделяют верхний и нижний червячки от полушарий
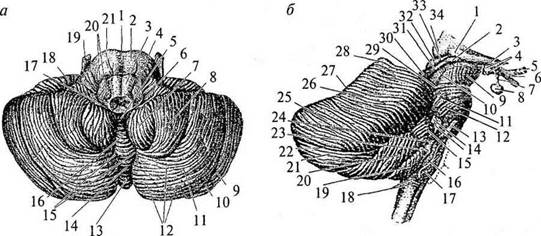
Рис. 30. Мозжечок:
вид снизу (о): 1 – борозда основной артерии; 2 – Варолиев мост; 3 – пирамида продолговатого мозга; 4 – олива; 5 – узелок червя; 6 – сосудистое сплетение IV желудочка; 7 – миндалина мозжечка; 8 – двубрюшная долька мозжечка; 9 – верхняя полулунная долька мозжечка; 10 – горизонтальная борозда мозжечка; 11 – нижняя полулунная долька; 12 – извилины мозжечка; 13 – бугор червя; 14 – пирамида червя; 15 – долинка мозжечка; 16 – язычок червя; 17 – ножка клочка; 18 – клочок; 19 – тройничный нерв; 20 – корешки языкоглоточного и блуждающего нервов; 21 – отводящий нерв; вид справа (б): 1 – латеральное коленчатое тело; 2 – медиальное коленчатое тело; 3 – зрительный тракт; 4 – мамиллярные тела; 5 – зрительный нерв; 6 – хиазма; 7 – воронка; 8 – гипофиз; 9 – ножка мозга; 10 – латеральная борозда среднего мозга; 11 – тройничный нерв; 12 – косой пучок моста; 13 – отводящий нерв; 14 – слуховой и лицевой нервы; 15 – клочок; 16 – олива; 17 – подъязычный нерв; 18 – наружные дугообразные волокна; 19 – миндалина мозжечка; 20 – двубрюшная долька мозжечка; 21 – нижняя полулунная долька мозжечка; 22 – горизонтальная борозда мозжечка; 23 – блуждающий нерв; 24 – языкоглоточный нерв; 25 – верхняя полулунная долька; 26 – четырехугольная долька; 27 – скат; 28 – вершина; 29 – борозда блокового нерва; 30 – нижние бугры четверохолмия; 31 – нижние ручки четверохолмия; 32 – верхние бугры четверохолмия; 33 .– верхние ручки четверохолмия; 34 – подушка мозжечка. На верхнем и нижнем червячках различают доли, которые состоят из нескольких извилин (рис. 29, 3).
Верхний червячок спереди назад состоит из следующих долей:
1) язычок мозжечка (рис. 30а, 16);
2) центральная долька (рис. 33, 23);
3) бугор (рис. 30а, 13);
4) лист червя, в виде очень узкой дольки располагающейся сзади, на границе перехода верхнего червячка в нижний (рис. 29, 3).
На нижнем червячке, в направлении спереди назад, различают следующие доли:
1) узелок (рис. 33, 20, к передним отделам которого присоединяется задний мозговой парус;
2) втулочка червя;
3) пирамида червя (рис. 33, 16).
На верхней поверхности полушарий мозжечка отмечают следующие борозды и дольки.
Четырехугольная долька (lobulus quadrangularis) (рис. 30б, 26) разделяется передней верхней бороздой (sulcus superior anterior) на переднюю и заднюю части. Четырехугольная долька ограничивается задней верхней бороздой от верхней полулунной дольки (lobulus semilunaris superior) (рис. 306, 25).
Впереди от lobulus quadrangularis залегают небольшие извилины, так называемые крылья центральной дольки. Снизу и спереди от последних лежат небольшие участки мозжечка – связи язычка (vinculo lingulae).
На нижней поверхности полушарий мозжечка располагаются следующие борозды и дольки.
Группа концентрически расположенных извилин образует миндалину (tonsilla) (рис. 30а, 7; 30б, 19). Снаружи и сзади от миндалины залегает двубрюшная долька (l o bulus bivénter) (рис. 30а, 8, 30б, 20). Двубрюшная долька соответствует пирамиде червячка.
Снаружи от миндалины и спереди от двубрюшной дольки располагается наиболее выступающая спереди часть нижней поверхности мозжечка – клочок (flocculus) (рис. 30а, 18; 306, 15).
Сзади от двубрюшной дольки лежит нижняя полулунная долька (lobulus semilunaris inferior) (рис. 30а, 11; 30б, 21).
Нейронная организация мозжечка существенно отличается от таковой стволовых структур. Основная масса нейронов сосредоточена на поверхности и создает кору мозжечка (cortex cerebelli). Площадь ее велика, так как кора имеется и на боковых поверхностях борозд (около 80%).
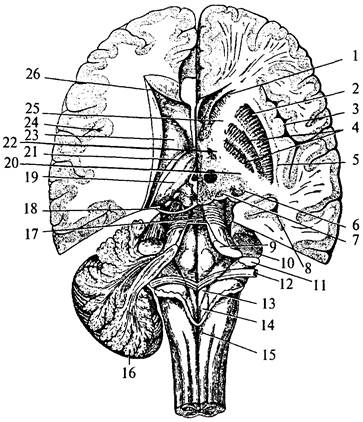
Притом что масса мозжечка составляет лишь 1/9 массы обоих больших полушарий, площадь поверхности его коры равна площади поверхности одного из них. Серое вещество коры, расположенное на поверхности ветвящихся борозд, пронизывает, как дерево, белое вещество. Поэтому рисунок, образуемый серым и белым веществом на срезах мозжечка, называют древом жизни мозжечка. В глубине белого вещества имеются скопления серого вещества – парные ядра мозжечка (nucleus cerebelli). B черве по обеим сторонам от средней линии имеются два ядра шатра (nucleus fastigii cerebélli) (рис. 31, 1), латералънее ядер шатра в полушариях мозжечка наблюдаем шаровидные ядра (nucleus globusus cerébelli) (рис. 31, 2). Латералънее последних, в полушариях имеются пробковидные ядра (nucleus emboliformis) (рис. 31, 3), а еще дальше – самые крупные ядра полушарий, зубчатые (nucleus dentatus) (рис. 31, 4), представляющие собой волнообразно изгибающуюся пластинку серого вещества.
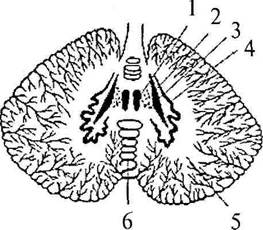
| Рис. 31. Ядра мозжечка: 1 – ядро шатра; 2 – шаровидное ядро; 3 – пробковидное ядро; 4 – зубчатое ядро; 5 – полушария мозжечка; 6 – червь мозжечка |
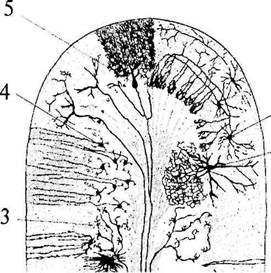
| Рис. 32. Схема строения коры мозжечка: 1 – звездчатая клетка; 2 – клетка Гольджи; 3 – моховидные волокна; 4 – клетки-зерна; 5 – клетка Пуркинье |
Кора мозжечка четко разделена на три слоя (рис. 32):
1) наружный – молекулярный слой (stratum moleculare); В нем находятся аксоны и дендриты клеток нижележащих cлоев, а также звездчатые и корзинчатые клетки (рис. 32, 1).
2) средний – ганглиозный слой (stratum ganglionaris);
Образован крупными грушевидными клетками Пуркинье, имеющими мощное, сильно ветвящееся дендритное дерево в молекулярном слое (рис. 32, 5).
3) внутренний – зернистый слой (stratum granulosum).
Аксоны клеток-зерен направляются в молекулярный слой, где Т-образно ветвятся и вступают в синаптические контакты с дендритами клеток Пуркинье, корзинчатых и звездчатых клеток (рис. 32, 4).
Дендритное дерево клеток Пуркинье располагается в плоскости, перпендикулярной оси борозды, а аксоны клеток-зерен – параллельно ей. На одну клетку Пуркинье приходится около 5 тыс. клеток-зерен. На соме и дендритах клеток Пуркинье заканчиваются еще и аксоны звездчатых и корзинчатых клеток, а также так называемых лазающих волокон от ядер оливы (пришедших к коре мозжечка по оливо-мозжечковым путям). Остальные афферентные пути заканчиваются в коре мозжечка в виде моховидных волокон (рис. 32, 5) на клетках-зернах, а также на звездчатых и корзинчатых клетках. Эфферентные выходы из коры мозжечка созданы аксонами клеток Пуркинье, заканчивающимися на клетках подкорковых ядер мозжечка. Из аксонов клеток ядер мозжечка состоят эфференты мозжечка, связывающие его с другими отделами ЦНС.
Афферентные и эфферентные волокна образуют в совокупности три пары мозжечковых ножек. Через нижнюю пару ножек мозжечок получает афференты от дорсального спиномозжечкового пути Флексинга, здесь проходят оливо-мозжечковый путь, пути от вестибулярных ядер VIIIпары нервов и ядер V, VII, IX и X пар черепно-мозговых нервов, а также от ядер Голля и Бурдаха продолговатого мозга. Через нижние ножки выходит лишь один эфферентный путь от ядер шатра к вестибулярным ядрам продолговатого мозга. Средние ножки имеют только афферентные волокна, идущие от собственных ядер моста, а также коллатерали от кортикоспинальных путей. Через эти ножки различные отделы коры больших полушарий (лобные, височные и затылочные) связаны с мозжечком, так как на собственных ядрах моста заканчиваются корково-мостовые нисходящие пути. Через верхние ножки мозжечок получает афферентные волокна от вентрального спиномозжечкового пути Говерса, а также от передних бугров четверохолмия. Основную же массу передних ножек составляют эфферентные волокна, идущие к красному ядру, ретикулярным ядрам и буграм четверохолмия среднего мозга, к таламическим и гипоталамическим ядрам промежуточного мозга. Через таламические ядра мозжечок связан с корой больших полушарий, а через красные ядра, ядра ретикулярной формации и вестибулярные ядра – со спинным мозгом.
Филогенез.
Филогенез.В ряду позвоночных животных наиболее примитивным является мозжечок круглоротых. Он представляет собой пластинку с наружным слоем волокон и внутренним клеточным слоем, которая связана с органом боковой линии и с вестибулярными ядрами. В дальнейшем развитие мозжечка было определено совершенствованием вестибуломозжечковых связей. Поэтому у рыб мозжечок развивается так, что становится высшей интегративной структурой. Мозжечок рыб состоит из тела и двух небольших возвышений. Возвышения имеются уже у круглоротых. Их называют древним мозжечком, а тело – старым мозжечком. При выходе животных на сушу мозжечок вначале редуцируется (у амфибий), а потом вновь развивается в мощную мозговую структуру (у рептилий и птиц). Развитие идет по пути совершенствования связей спинного мозга с мозжечком и ослабления связей с вестибулярной системой. У высших рептилий (крокодилов) и птиц формируется кора мозжечка, имеющая два слоя клеток: гранулярный и молекулярный (с клетками Пуркинье). Тело мозжечка делится на три доли: переднюю, среднюю и заднюю. У млекопитающих в мозжечке возникают структуры, тесно связанные с корой больших полушарий – новый мозжечок. Впервые появляются полушария мозжечка, а также его средние ножки. Они образуются из тела мозжечка низших позвоночных. У грызунов в мозжечке появляются три подкорковых ядра (шатра, зубчатое и срединное). И только у приматов срединное ядро распадается на шаровидное и пробковидное. Наибольшего развития достигает зубчатое ядро.
Развитый мозжечок высших млекопитающих состоит из трех частей: древний (paleocerebellum) контролирует вестибулярную функцию (его каудальные отделы); старый (archicerebellum) связан со спинным мозгом (передняя доля) и новый (neocerebellum) – с корой больших полушарий.
Онтогенез.
Онтогенез.В онтогенезе мозжечок развивается из мозжечковой пластинки, являющейся крышей заднемозгового пузыря (metencephaton). Полушария мозжечка образуются из боковых частей, а из средней части этой пластинки формируется червь.
Средний мозг
Средний мозг (mesencephalon) является верхней частью мозгового ствола. Средний мозг делят на дорсальную часть – крышу мозга (tectum) и вентральную – ножки мозга (pedunculi cerebri) (рис. 33, 26; 34, 5). Полость среднего мозга представлена узким каналом – Сильвиевым водопроводом (aqueductus cerebri), который соединяет III и IV мозговые желудочки (рис. 34, 1).
Крыша среднего мозга, или пластинка четверохолмия (рис. 33, 11) образована двумя верхними (colliculi superior) (см. рис. 36, 8а) и двумя
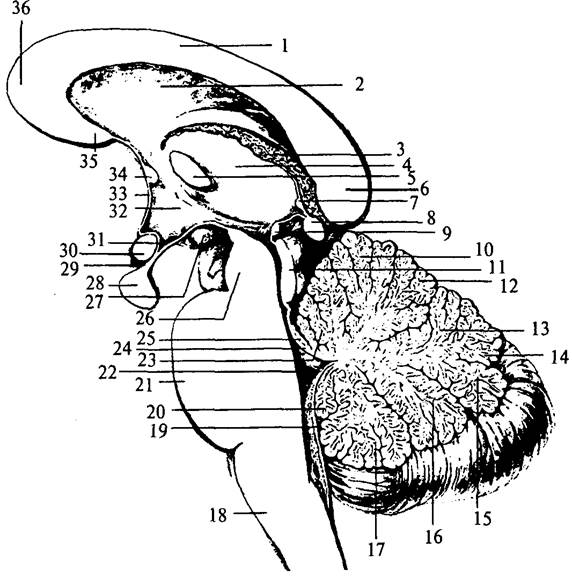
Рис. 33. Сагиттальный срез ствола мозга:
1 – ствол мозолистого тела; 2 – прозрачная перегородка; 3 – сосудистое сплетение III желудочка; 4 – таламус; 5 – межталамическое сращение; 6 – валик мозолистого тела; 7 – комиссура поводка; 8 – шишковидная железа; 9 – задняя комиссура; 10 – вершина мозжечка; 11 – четверохолмие; 12 – скат; 13 – червь мозжечка; 14 – листок червя; 15 – бугор червя; 16 – пирамида червя; 17 – язычок червя; 18 – продолговатый мозг; 19 – нижний мозговой парус; 20 – узелок; 21 – Варолиев мост; 22 – полость IV желудочка; 23 – центральная долька; 24 – верхний мозговой парус; 25 – язычок мозжечка; 26 – ножки среднего мозга; 27 – мамиллярное тело; 28 – гипофиз; 29 – воронка; 30 – хиазма; 31 – углубление воронки; 32 – гипоталамус; 33 – конечная пластинка; 34 – передняя комиссура; 35 – клюв мозолистого тела; 36 – колено мозолистого тела нижними холмиками (colliculi inferior) (рис. 36, 8б). От каждой пары холмиков в направлении промежуточного мозга отходят проводящие пути – пары ручек холмиков (branchii colliculus) (рис. 305, 31, 33). Ручки верхних холмиков заканчиваются в латеральных коленчатых телах (рис. 34, 15), а нижних – в медиальных коленчатых телах промежуточного мозга (рис. 34, 76).
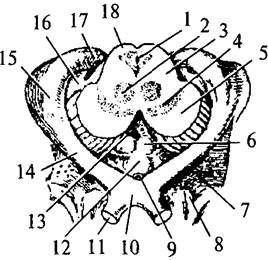
| Рис. 34. Разрез среднего мозга (изображены примыкающие к нему части промежуточного мозга): 1 – Сильвиев водопровод; 2 – красное ядро; 3 – покрышка среднего мозга; 4 – черная субстанция; 5 – ножки среднего мозга; 6 – сосцевидные тела; 7 – переднее продырявленное пространство; 8 – обонятельный треугольник; 9 – воронка; 10 – зрительный перекрест; 11 – зрительный нерв; 12 –серый бугор; 13 – заднее продырявленное пространство; 14 – зрительный тракт; 15 – латеральное коленчатое тело; 16 – медиальное коленчатое тело; 17 – подушка таламуса; 18 – крыша среднего мозга |
На основании мозга, впереди моста лежат ножки мозга – два симметричных толстых расходящихся валика, которые упираются в большие полушария (рис. 34, 5). Между ножками имеется межножковая ямка (fossa interpeduncularis), закрытая задним продырявленным пространством (substantia perforata posterior) (рис. 34, 13). На медиальной поверхности каждой ножки выходят волокна III пары глазодвигательного нерва (III – п. oculomotorius). Волокна IV пары блокового нерва (IV– п. trochlearis) отходят от-дорсальной поверхности среднего мозга. Оба нерва среднего мозга являются двигательными (рис. 26; 27; 35, 6).
На поперечном срезе среднего мозга выделяют три отдела:
1) крыша среднего мозга (tectum mesencephali) (рис. 35, 2);
2) покрышка (tegmentum mesencephali) (рис. 34, 3);
3) основание ножек мозга (basis pedunculi cerebralis) (рис. 35, 5).
Наружная поверхность крыши среднего мозга покрыта тонким слоем белого вещества, переходящим в ручки холмиков. Под этим слоем находятся ядра верхних (nucleus colliculi superioris) и нижних (nucleus colliculi inferioris) бугров четверохолмия. Ядра верхних бугров имеют слоистое строение. К ним приходят афферентные волокна от зрительного тракта, от спинного мозга по спинотектальным путям, а также коллатерали от латеральной имедиальной петель. Отходят эфферентные волокна к двигательным ядрам ствола и спинного мозга по тектобульбарным и тектоспинальным путям. Верхними ручками передние бугры связаны с латеральными коленчатыми телами. В ядрах нижних бугров заканчивается часть волокон латеральной петли. Эфферентными же волокнами они сплетаются с медиальными коленчатыми телами (по нижним ручкам), а также со спинным мозгом и мозговым стволом (по тектоспинальным и тектобульбарным путям).
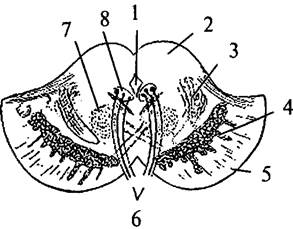
| Рис. 35. Поперечный разрез через ножки мозга: 1 – Сильвиев водопровод; 2 – крыша среднего мозга; 3 – медиальная петля; 4 – черная субстанция; 5 – основание ножек; 6 – глазодвигательный нерв; 7 – красное ядро; 8 – ядро глазодвигательного нерва |
Верхнее двухолмие является подкорковым зрительным центром, а нижнее двухолмие служит местом переключения слуховых путей и играет роль слухового подкоркового центра.
В покрышке среднего мозга имеются красные ядра (nucleus ruber) (рис. 34, 2; 35, 7), которые дают начало руброспинальному пути. В красных ядрах заканчиваются волокна верхних ножек мозжечка. Вокруг Сильвиева водопровода (рис. 34, 1) располагается центральное серое вещество (substantia grisea centralis). В нем находятся ядра ретикулярной формации среднего мозга, получающие коллатерали от проходящих здесь восходящих и нисходящих путей, а свои длинные аксоны направляющие к другим мозговым структурам и к коре больших полушарий. Ядра блокового нерва (IV пара) лежат в центральной части серого вещества, непосредственно у Сильвиева водопровода, на уровне нижних бугров четверохолмия. Под дном водопровода, на уровне верхних бугров четверохолмия, находятся ядра глазодвигательных нервов (III пара) (рис. 35, 5). Латерально и кверху от красных ядер находятся слои медиальных петель, идущих от покрышки моста.
Между покрышкой и основанием ножек располагается ядро, состоящее из клеток, богатых меланином, – черная субстанция (substantia nigra) (см. рис. 35, 4).
Основание ножек мозга лишено ядер и образовано нисходящими из коры больших полушарий корковоспинномозговыми, корковомостовыми путями.
Средний мозг является первичным зрительным и слуховым центром, осуществляющим быстрые рефлекторные реакции (оборонительные и ориентировочные). Кроме того, красные ядра и черная субстанция являются ядрами, контролирующими тонус мускулатуры и движения.
Филогенез.
Филогенез.У круглоротых средний мозг представлен крышей, имеющей связи с продолговатым мозгом. У ганоидных и костистых рыб развивается покрышка, где формируются ядра черепно-мозговых нервов, появляются связи с мозжечком. У амфибий развита крыша, представленная двухолмием. У рептилий возникает красное ядро и формируется руброспинальный путь. У млекопитающих появляется черная субстанция, крыша из двухолмия превращается в четверохолмие с сохранением зрительной функции за передним двухолмием и формированием слуховой функции в связи с задним двухолмием. Кроме того, устанавливаются связи среднего мозга с таламусом, базальными ганглиями и корой больших полушарий. В центральной части среднего мозга развивается массивная ретикулярная формация с множественными восходящими и нисходящими путями.
Онтогенез.
Онтогенез.В онтогенезе средний мозг формируется из среднего мозгового пузыря, стенки которого равномерно утолщаются, превращая его полость в узкий канал – водопровод.
Промежуточный мозг
Промежуточный мозг (diencéphalon) каудально соединяется со средним мозгом, а рострально переходит в большие полушария конечного мозга. Полость промежуточного мозга представляет собой вертикальную щель, расположенную в серединной сагиттальной плоскости, это IIIмозговой желудочек (ventriculus tertius) (рис. 36, 11). Сзади он переходит в водопровод среднего мозга, а впереди соединяется с двумя боковыми желудочками больших полушарий посредством двух межжелудочковых отверстий Монро (forâmena interventricularià) (см. рис. 40, 16). Боковые стенки IIIжелудочка образованы медиальными поверхностями правого и левого таламусов, дно – гипоталамусом и субталамусом. Передняя граница подходит к нисходящим колоннам свода (columnae fornicis) (рис. 36, 4), ниже к передней мозговой комиссуре (comissura anterior) (рис. 36, 5) и далее к конечной пластинке (lamina terminalis). Задняя стенка состоит из задней комиссуры (comissura posterior) над входом в водопровод мозга (рис. 36, 7). Крыша III желудочка состоит из эпителиальной пластинки. Над ней располагается сосудистое сплетение. Выше сплетения проходит свод, а еще выше – мозолистое тело (см. рис. 33, 1). По боковым стенкам III желудочка от межжелудочковых отверстий до входа в водопровод мозга проходят гипоталамические борозды, отделяющие таламусы от гипоталамуса. Таламусы соединяются между собой в средней части III желудочка спайкой – межталамическим сращением (adhesio interthalamica) (см. рис. 33, 5; 36, 6). Промежуточный мозг включает в себя несколько структур: собственно зрительный бугор – таламус, метаталамус, гипоталамус, субталамус, эпиталамус, гипофиз.
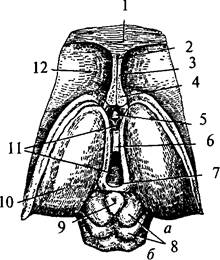
| Рис. 36. Промежуточный и средний мозг (вид сверху): 1 – мозолистое тело; 2 – полость прозрачной перегородки; 3 – прозрачная перегородка; 4 – свод (поперечный разрез передних ножек); 5 – передняя комиссура; 6 – межталамическое сращение; 7 – задняя комиссура; 8 – бугры четверохолмия (а – верхние, б – нижние); 9 – шишковидная железа; 10 –таламус; 11 – III желудочек; 12 – хвостатое ядро |
Таламус
Таламус (thalamus) – основная часть промежуточного мозга. Он составляет боковые стенки III желудочка (рис. 36, 70). Включает в себя собственно зрительный бугор и метаталамус (латеральные и медиальные коленчатые тела). Форма таламуса яйцевидная, узкая часть направлена назад. Выступающая задняя часть таламуса называется подушкой (pulvinar), а в передней части таламус имеет передний бугорок. Ниже и латеральнее подушки располагаются продолговато-овальные бугорки: медиальное (corpus geniculatum mediale) и латеральное (corpus geniculatum laterale) коленчатые тела (см. рис. 305, 1, 2; 34, 15, 16). Медиальная поверхность таламуса образует боковую стенку III желудочка, верхняя и латеральная прилегают к внутренней капсуле больших полушарий, а нижняя граничит с гипоталамусом (см. рис. 42, 7).
Метаталамус (metathalamus) представлен коленчатыми телами, расположенными ниже и латеральнее подушки. Медиальное коленчатое тело выражено лучше, лежит под подушкой зрительного бугра и наряду с нижними буграми четверохолмия является подкорковым центром слуха (рис. 34, 16). Латеральное коленчатое тело – небольшое возвышение, лежащее на нижнелатеральной поверхности подушки. Оно вместе с верхними буграми четверохолмия является подкорковым зрительным центром (см. рис. 34, 75).
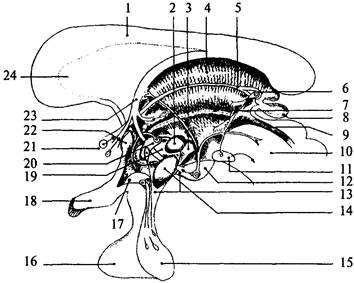
| Рис. 37. Схема основных ядерных групп промежуточного мозга: 1 – мозолистое тело; 2 – дорсомедиальное ядро гипоталамуса; 3 – заднее ядро гипоталамуса; 4 – дорсальные отделы таламуса; 5 – мозговая полоска зрительного бугра; 6 – уздечка; 7 – гипоталамическая борозда; 8 – эпифиз; 9 – Сильвиев водопровод; 10 – средний мозг; 11 – межножковое ядро; 12 – сосцевидное тело; 13 – серый бугор; 14 – вентромедиальное ядро гипоталамуса; 15 – нейрогипофиз; 16 – аденогипофиз; 17 – супраоптическое ядро гипоталамуса; 18 – хиазма; 19 – латеральное ядро гипоталамуса; 20 – паравентрикулярное ядро гипоталамуса; 21 – ядро прозрачной перегородки; 22 – передняя спайка; 23 – свод; 24 – прозрачная перегородка |
В подушке и коленчатых телах находятся одноименные ядра. В наружные коленчатые тела входят так называемые зрительные тракты, которые являются зрительными путями, составленными уже перекрещенными аксонами ганглиозных клеток сетчатки. Внутренняя структура таламуса представляет собой ядерные скопления серого вещества, разделенного белым веществом. В таламусе имеется около 150 ядер (рис. 37). Их подразделяют на шесть групп: передняя, средней линии, медиальная, латеральная, задняя и претектальная.
В соответствии с функциями различают специфические и неспецифические ядра таламуса. Специфические, в свою очередь, представляют собой переключательные (сенсорные и несенсорные) и ассоциативные ядра. Аксоны клеток ядер таламуса подходят к определенным участкам коры. Переключательные ядра получают афференты от разных сенсорных систем или от других отделов мозга, а свои афференты направляют к определенным проекционным зонам коры. В ассоциативных ядрах заканчиваются афференты от других таламических ядер, а аксоны их клеток идут к ассоциативным зонам коры.
Неспецифические ядра не имеют специфических афферентных связей с отдельными сенсорными системами, а их афференты устремляются диффузно ко многим участкам коры. Переключательные ядра зрительной и слуховой сенсорных систем – ядра латерального и медиального коленчатых тел, а соматосенсорной системы – заднее вентральное ядро таламуса. Ассоциативными ядрами являются латеральные и медиальные ядра подушки. Неспецифические ядра сосредоточены преимущественно в латеральной, медиальной и средней группах ядер таламуса. Таламус связан со всеми отделами ЦНС. Таламус участвует в переработке сенсорных стимулов, идущих к коре больших полушарий, а также регулирует цикл бодрствование – сон.
Гипоталамус
Гипоталамус (hypothalamus) – небольшое образование передне-нижней части промежуточного мозга, весом около 4 г (см. рис. 33, 32). Он образует дно III желудочка и отделяется от таламусов гипоталамическими бороздами (рис. 37, 7). На основании мозга между его ножками и сзади от перекреста зрительных нервов (хиазмы) видны основные структуры гипоталамуса. Серый бугор (tuber cineréum) расположен между сосцевидными телами и перекрестом зрительных нервов, с боков ограничен зрительными трактами (см. рис. 34, 12). Он соединен с воронкой (infundibulum) (см. рис. 34, 9), которая переходит вентральнее в ножку гипофиза и далее в гипофиз (hypophysis). За серым бугром следуют белого цвета парные круглые образования – сосцевидные тела (corpora mamillaria) (см. рис. 34, 6). Своей передней частью гипоталамус прилежит к конечной пластинке мозга, а передненижней – к зрительному перекресту. Сосцевидные тела граничат с задним продырявленным пространством (substantia perforata posterior). Эта часть поверхности мозга называется продырявленным пространством, потому что пронизана многочисленными сосудами. Серое вещество гипоталамуса образует ядра, которые подразделяются на пять групп: преоптическую, переднюю, среднюю, наружную и заднюю группы (рис. 37; 38).
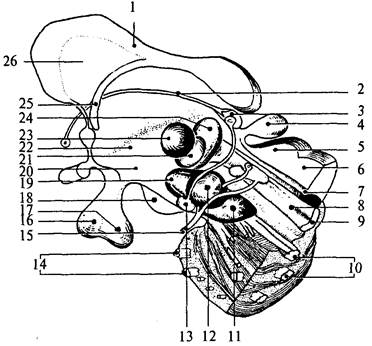
| Рис. 38. Схема основных ядерных групп промежуточного мозга и среднего мозга: 1 – мозолистое тело; 2 – мозговая полоска зрительного бугра; 3 – уздечка; 4 – эпифиз; 5 – верхний бугорок четверохолмия; 6 – нижний бугорок четверохолмия; 7 – Сильвиев водопровод; 8 – медиальный продольный пучок; 9 – медиальная петля; 10 – пирамидный тракт; 11 – черное вещество среднего мозга; 12– красное ядро; 13 – межножковое ядро; 14 – поперечные волокна моста; 15 – глазодвигательный нерв; 16 – аденогипофиз; 17 – нейрогипофиз; 18 – сосцевидное тело; 19 – субталамическое ядро; 20 – гипоталамус; 21 – вентропостеромедиальное ядро таламуса; 22 – гипоталамическая борозда; 23 – центральное медиальное ядро таламуса; 24 – вентропостеролатеральное ядро таламуса; 25 – свод; 26 – прозрачная перегородка |
Преоптическая группа включает медиальное и латеральное преоптические ядра, а также перивентрикулярное ядро. В переднюю группу входят: супраоптическое, супрахиазматическое и околожелудочковое ядра. Средняя группа – это вентромедиальные и дорсомедиальные ядра серого бугра. К наружной группе относят скопление серого вещества – латеральное гипоталамическое поле. Ядра задней группы – заднее гипоталамическое, латеральные и медиальные ядра сосцевидных тел: латеральное мамиллярное ядро меньше медиального.
Ядра гипоталамуса тесно связаны с другими мозговыми структурами. Так, ядра сосцевидных тел получают большое количество волокон из гиппокампа (структуры конечного мозга) через свод (fornix) и из покрышки среднего мозга через мамиллярную ножку.
Эфферентные волокна сосцевидных тел образуют их главный пучок (tractus principalis), или мамиллоталамический (Вик д'Азира), который направляется к переднему ядру таламуса, а мамиллосегментальный – к покрышке среднего мозга (в пучке Вик д'Азира проходит 0,7 млн волокон). Ядра боковых и средних групп гипоталамуса имеют двусторонние связи с базальными ядрами больших полушарий головного мозга. Ядра серого бугра получают афференты от обонятельного мозга, а передняя группа ядер – от свода. Эфференты ядер гипоталамуса связывают их с таламусом, субталамусом и другими подкорковыми структурами. Особо следует выделить связи передней группы ядер с задней долей гипофиза (нейрогипофизом). Аксоны клеток этих ядер создают мощный пучок волокон, проходящих в ножке гипофиза (около 100 тыс. волокон образуют аксоны клеток супраоптического ядра).
В гипоталамусе следует различать три основные группы нейросекреторных клеток: 1) пептидергические; 2) либерин- и статинергические; 3) моноаминергические. Однако это разделение весьма условно, так как одни и те же клетки могут синтезировать два типа нейрогормонов.
Паравентрикулярное и супраоптическое ядра связаны с нейрогипофизом путем прорастания в него аксонов нервных клеток, образующих эти ядра и формирующих гипоталамо-нейрогипофизарную систему. В супраоптическом и паравентрикулярном ядрах синтезируются два пептидных гормона, секретирующихся из нейрогипофиза. Это вазопрессин и окситоцин.
Гипоталамус является высшим подкорковым центром интеграции нервных и эндокринных влияний, вегетативных и эмоциональных компонентов поведенческих реакций и тем самым обеспечивает регуляцию постоянства внутренней среды.
Субталамус
Сзади от гипоталамуса, в задненижнем участке промежуточного мозга, в межножковой его области над задним продырявленным пространством находится субталамус (subthalamus). В этой части промежуточного мозга у человека располагается субталамическое ядро Люиса (nucleus subthalamicus Luisi) (рис. 38, 19).
Эпиталамус
По границам эпителиальной пластинки и боковым стенкам III желудочка протянулись мозговые полоски, расширяющиеся сзади в треугольники поводка (trigonum habenularae). B этих треугольниках серое вещество формирует ядра поводка (nucleus habenulae). Треугольники поводка направляются к середине дорсальной поверхности среднего мозга и переходят в поводки (habenula). Соединяясь, поводки образуют спайку, к задней поверхности которой прикреплено шишковидное тело (corpus pineale), или эпифиз (epiphysis) (см. рис. 33, 8; 36, 9; 37, 8; 38, 4).
Эпифиз является железой внутренней секреции. Мозговые полоски, треугольники, поводки и эпифиз образуют эпиталамус (см. рис. 31, 34, 35). Эпифиз является железой внутренней секреции, контролирующей течение циркадианных ритмов и тормозящей активность половых желез до наступления периода полового созревания.
Гипофиз
Гипофиз (hypophysis), или нижний мозговой придаток, располагается на вентральной поверхности мозга в основании черепа в ямке турецкого седла (см. рис. 33, 28; 37, 15, 16; 38, 16, 17). После рождения, по мере роста и развития ребенка вес гипофиза увеличивается и к 14 годам достигает массы гипофиза взрослого человека: 0,7 г у девушек и 0,6 г у юношей. В период беременности масса гипофиза у женщин увеличивается до 1 г. Гипоталамус и гипофиз составляют единую функциональную систему, обеспечивающую совместную регуляцию функций нервным и гуморальным путем.
По своему строению и эмбриогенезу гипофиз не однороден. В гипофизе различают две главные части: нейрогипофиз и аденогипофиз, имеющие различное эмбриональное происхождение и строение.
Нейрогипофиз
Нейрогипофиз (см. рис. 37, 15; 38, 17) представляет собой производное дна воронки промежуточного мозга. Он находится в тесной морфологической и функциональной связи с гипоталамусом, в нем заканчиваются волокна гипоталамо-гипофизарного тракта, идущего от супраоптического и паравентрикулярного ядер гипоталамуса (см. рис. 37, 17, 20).
Аденогипофиз (передняя доля)
Аденогипофиз (передняя доля) развивается из эпителиального выпячивания (кармана Ратке) крыши кишечной трубки (см. рис. 37, 16; 38, 16). Передняя доля гипофиза имеет тесную сосудистую связь с гипоталамусом. Здесь артерии ветвятся на капилляры, образуя плотное сплетение в форме мантии на поверхности срединного возвышения. Капиллярные ветви этого сплетения образуют вены, достигающие передней доли гипофиза, здесь вены вновь распадаются на капилляры, пронизывающие всю долю. Вся эта сложная система кровеносных сосудов носит название портальной. По ней в аденогипофиз из гипоталамуса поступают пептидные гормоны (либерины и статины), регулирующие синтез и секрецию гормонов аденогипофиза. Нейрогипофиз имеет собственную, не зависящую от портальной системы, систему кровоснабжения.
В аденогипофизе секретируется два типа гормонов – эффекторные, т.е. реализующие свои свойства непосредственно в организме, и тропные – оказывающие регулирующее влияние на периферические железы внутренней секреции. Всего в аденогипофизе синтезируется шесть гормонов – гормон роста, пролактин, тиреотропин, адренокортикотропный гормон (АКТГ), фолликулостимулирующий гормон, лютенизирующий гормон. Фолликулостимулирующий и лютенизирующий гормоны объединяются в группу гонадотропных гормонов.
За последние годы было установлено, что практически все биологически активные вещества, секретируемые нейронами гипоталамо-гипофизарной системы, имеют пептидную природу.
Филогенез.
Филогенез.Вфилогенезе структуры промежуточного мозга появляются и развиваются на разных его этапах. У низших позвоночных животных уже есть некоторые эпиталамические, метаталамические и гипоталамические структуры. У круглоротых таламус уже достаточно выражен и разделяется на дорсальную и вентральную части, включающие в себя несколько ядер. Гипоталамус является также выраженной структурой. У костистых рыб происходит усложнение в организации таламуса и дифференциация ряда ядерных групп. У амфибий и рептилий развиваются зрительные бугры. Они становятся специальными структурами и связывают средний мозг с конечным. У рептилий промежуточный мозг выполняет вместе с некоторыми структурами конечного мозга высшие интегративные функции. Дорсальный таламус у них отделяется от вентрального, на этом этапе выделяются четкие ядра с характерным строением и связями. В дорсальном таламусе их насчитывается девять, в вентральном – семь. В переднем отделе гипоталамуса у рептилий появляются зачатки супраоптического и паравентрикулярного ядер.
У птиц дифференциация ядер гипоталамуса хорошо выражена, тогда как таламус усложняется незначительно. Особенно бурно идет развитие дорсального таламуса у млекопитающих, так как он выполняет функцию основного коллектора сенсорных путей к коре больших полушарий. У млекопитающих наряду с развитием релейных (переключательных) ядер таламуса развиваются ассоциативные ядра. Происходит усиленное развитие таламуса и формируется таламонеокортикальная система интеграции.
Онтогенез.
Онтогенез.В онтогенезе структуры промежуточного мозга возникают из стенок второго мозгового пузыря (diencephalon) пятипузырной стадии развития головного мозга. Глазные пузырьки превращаются в глазные бокалы и глазные стебельки. Позже из них строятся сетчатка глаза и зрительные проводящие пути. Из боковых стенок второго пузыря формируются структуры дорсального (thalamus) и вентрального (subthalamus) таламусов. Верхней стенкой являются эпиталамические структуры, нижней – гипоталамус.
Из верхней части задней стенки развиваются забугорные структуры (metathalamus). B этой части мозговой трубки все структуры образуются во время онтогенеза из крыловидной пластинки (пограничная борозда и основная пластинка заканчиваются на уровне среднемозгового пузыря). Поэтому здесь не возникают ни моторные (двигательные), ни вегетативные ядра, а следовательно, нет и соответствующих черепно-мозговых нервов. Все ядра промежуточного мозга являются либо сенсорными (переключательными), либо интегративными (ассоциативными). Имеются здесь и неспецифические ядра.
Гипоталамус отделяется от соседних участков мозга на 2-м месяце внутриутробной жизни. Вслед за этим начинается формирование шести гипоталамических ядер, которые представляют собой скопление нейронов с определенными функциями. Дифференцировка входящих в них клеток продолжается до 6-го месяца внутриутробной жизни, а завершается еще позднее. В четырех из шести ядер, вырабатываются гормоны, которые по системе сосудов направляются в аденогипофиз. Гипоталамо-аденогипофизарную систему составляют супрахиазматическое ядро, вентромедиальное, дорсомедиальное и аркуатное ядра. Сосудистая система появляется на 14-й неделе в виде первых капиллярных петель, и ее формирование завершается к моменту рождения. Синтезируемые в этих ядрах регуляторные пептиды обнаруживаются в аденогипофизе уже на 10-й неделе развития плода. Однако, по некоторым наблюдениям, в первые три месяца внутриутробной жизни, а возможно, до первой половины беременности гипофиз не подчиняется контролю гипоталамуса. Это объясняется незрелостью нейросекреторных клеток и недостаточным развитием портальной системы сосудов.
Вазопрессин появляется в гипофизе плода на 15–17-й, а окситоцин – на 18–19-й неделях внутриутробного развития. К 6-му месяцу беременности содержание их значительно возрастает. Уже в этот период они принимают участие в регуляции жизнедеятельности плода. Установление гипоталамического контроля над эндокринными железами происходит к концу внутриутробного развития.
Конечный мозг
Конечный мозг (telencephalon) – самый большой отдел головного мозга. Он состоит из двух полушарий (hemisphéria cerebri), которые разделены продольной мозговой щелью. Полушария сильно разрастаются и покрывают собой все остальные отделы головного мозга. Длина полушарий 170 мм, высота 120 мм.
Полушария вместе со всем головным мозгом покрыты тремя мозговыми оболочками (рис. 39). Твердая мозговая оболочка складкой проникает глубоко между правым и левым полушариями и между затылочным полюсом полушарий и мозжечком. Паутинная оболочка входит во все борозды на поверхности. Под паутинной оболочкой находится подпаутинное пространство, заполненное спинномозговой жидкостью. Расширения подпаутинного пространства называются цистернами мозга. Мягкая мозговая оболочка покрывает всю поверхность полушарий (см. рис. 39). Подпаутинное пространство составляет функционально единое целое с венозной системой мозга и системой его желудочков (рис. 40).
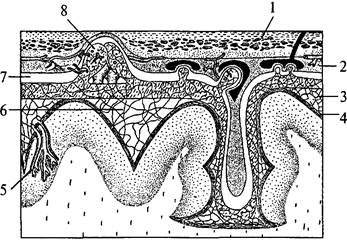
| Рис. 39. Схема взаимоотношений оболочек головного мозга: 1 – кость; 2 – твердая мозговая оболочка; 3 – паутинная оболочка; 4 – мягкая (сосудистая) оболочка; 5 – кровеносный сосуд мозга; 6 – подпаутинное пространство; 7 – субдуральное пространство; 8 – пахионовы грануляции |
| Рис. 40. Система желудочков и цистерн мозга: а – схема поверхности мозга: 1 – цистерны мозга; б – схема сагиттального среза мозга: 1 – листки твердой оболочки мозга; 2 – паутинная оболочка; 3 – мягкая оболочка; 4 – задний рог бокового желудочка; 5 – нижний рог бокового желудочка; 6 – грануляции паутинной оболочки; 7 – отверстие Мажанди; 8 – центральный канал; 9 – отверстие Люшке; 10 – сосудистое сплетение IV желудочка; 11 – IV желудочек; 12 – Сильвиев водопровод; 13 – III желудочек; 14 – сосудистое сплетение III желудочка; 15 – передний рог бокового желудочка; 16 – Монроево отверстие; 17 – сосудистое сплетение бокового желудочка; 18 – боковой желудочек; 19 – корковые ветви мозговой вены |
Каждое полушарие имеет свою щелевидную полость – боковой желудочек (рис. 40, 18). Условно желудочек левого полушария считается I, а правого – II. Вслед за ростом полушарий боковые желудочки разрастаются и принимают своеобразную форму. Кроме центральной части каждый желудочек имеет распространяющийся в лобную долю передний рог (рис. 40, 15), идущий в глубине затылочной доли задний рог (рис. 40, 4) и отходящий от него вниз и латерально нижний рог (рис. 40, 5), являющийся полостью височной доли. Боковые желудочки сообщаются с полостью IIIжелудочка и имеют в своих стенках сосудистое сплетение. Медиальной стенкой каждого переднего рога является тонкая прозрачная пластинка. Правая и левая пластинки образуют между передними рогами общую прозрачную перегородку (septum pellucidum) (см. рис. 33, 2; 36, 3; 38, 26).
Расположение белого и серого вещества в полушариях сходно с таковым в мозжечке: в глубине белого вещества расположены ядра серого вещества, а вся поверхность полушарий покрыта серым веществом.
Базальные ганглии
В основании больших полушарий (нижняя стенка боковых желудочков) расположены ядра серого вещества – базальные ганглии. Они составляют примерно 3% от объема полушарий. Все базальные ганглии функционально объединены в две системы. Первая группа ядер представляет собой стриопаллидарную систему (рис. 41, 42, 43). К ним относятся: хвостатое ядро (nucleus caudatus), скорлупа (putamen) и бледный шар (globus pallidus). Скорлупа и хвостатое ядро имеют слоистую структуру, и поэтому их общее название – полосатое тело (corpus striatum). Бледный шар не имеет слоистости и выглядит светлее стриатума. Скорлупа и бледный шар объединены в чечевицеобразное ядро (nucleus lentiförmis). Скорлупа образует наружный слой чечевицеобразного ядра, а бледный шар – внутренние его части. Бледный шар, в свою очередь, состоит из наружного и внутреннего члеников.
Анатомически хвостатое ядро тесно связано с боковым желудочком. Расположенная впереди и медиально расширенная его часть – головка хвостатого ядра образует боковую стенку переднего рога желудочка, тело ядра – нижнюю стенку центральной части желудочка, а тонкий хвост – верхнюю стенку нижнего рога. Следуя форме бокового желудочка, хвостатое ядро охватывает дугой чечевицеобразное ядро (рис. 42, 1; 43, 1/). Друг от друга хвостатое и чечевицеобразное ядра отделены прослойкой белого вещества – частью внутренней капсулы (capsula interna). Другая часть внутренней капсулы отделяет чечевицеобразное ядро от нижележащего таламуса (рис. 43, 4).
| Рис. 41. Полушария мозга на разных уровнях горизонтального разреза: (справа – ниже уровня дна бокового желудочка; слева – над дном бокового желудочка; IV желудочек мозга вскрыт сверху): 1 – головка хвостатого ядра; 2 – скорлупа; 3 – кора мозгового островка; 4 – бледный шар; 5 – ограда; 6 – хвост хвостатого ядра; 7 – ядра медиального коленчатого тела; 8 – нижний рог бокового желудочка; 9 – верхние ножки мозжечка; 10 – средние ножки мозжечка; 11 – нижние ножки мозжечка; 12 – мозговые полоски (волокна проводящих путей слуховой системы); 13 – треугольник подъязычного нерва; 14 – треугольник блуждающего нерва; 15 – задвижка; 16 – часть мозжечка; 17 – верхний мозговой парус; 18 – блоковый нерв; 19 – таламус; 20 – задняя ножка внутренней капсулы; 21 – конечная пластинка; 22 – колено внутренней капсулы; 23 – хвостатое ядро; 24 – кора мозгового островка; 25 – передняя ножка внутренней капсулы; 26 – передний рог бокового желудочка |
Таким образом, строение дна бокового желудочка (представляющего собой стриопаллидарную систему) схематично можно представить себе так: стенку самого желудочка образует слоистое хвостатое ядро, затем ниже идет прослойка белого вещества – внутренняя капсула, под ней слоистая скорлупа, еще ниже бледный шар и опять слой внутренней капсулы, лежащий на ядерной структуре промежуточного мозга – таламусе.
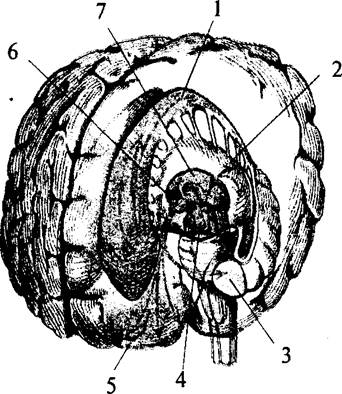
| Рис. 42. Топография базальных ядер конечного мозга и стволовых структур (вид слева спереди): 1 – хвостатое ядро; 2 – скорлупа; 3 – миндалина; 4 – черная субстанция; 5 – лобная кора; 6 – гипоталамус; 7 – таламус |
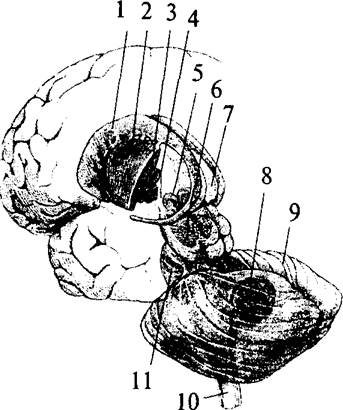
| Рис. 43. Топография базальных ядер конечного мозга и стволовых структур (вид слева сзади): 1 – хвостатое ядро; 2 – скорлупа; 3 – бледный шар; 4 – внутренняя капсула; 5 – субталамическое ядро; 6 – черная субстанция; 7 – таламус; 8 – подкорковые ядра мозжечка; 9 – мозжечок; 10 – спинной мозг; 11 – мост |
Стриопаллидарная система получает афферентные волокна от неспецифических медиальных таламических ядер, лобных отделов коры больших полушарий, коры мозжечка и черной субстанции среднего мозга. Основная масса эфферентных волокон стриатума радиальными пучками сходится к бледному шару. Таким образом, бледный шар является выходной структурой стриопаллидарной системы. Эфферентные волокна бледного шара идут к передним ядрам таламуса, которые связаны с фронтальной и теменной корой больших полушарий. Часть эфферентных волокон, не переключающихся в ядре бледного шара, идет к черной субстанции и красному ядру среднего мозга. Стриопаллидум (рис. 41; 42), совместно со своими проводящими путями, входит в экстрапирамидную систему,оказывающую тоническое влияние на моторную деятельность. Эта система контроля над движениями называется экстрапирамидной потому, что переключается на пути к спинному мозгу, минуя пирамиды продолговатого мозга. Стриопаллидарная система является высшим центром непроизвольных и автоматизированных движений, снижает мышечный тонус, тормозит движения, осуществляемые двигательной корой.
Латеральнее стриопаллидарной системы базальных ганглиев расположена тонкая пластинка серого вещества – ограда (claustrum). Она ограничена со всех сторон волокнами белого вещества – наружной капсулой (capsula externa).
Остальные базальные ядра входят в лимбическую систему мозга (см. раздел 6.2.5.3). Впереди от конца нижнего рога бокового желудочка в белом веществе височной доли больших полушарий расположена плотная группа ядер – миндалевидное тело (amigdalae) (рис. 42, 3). И наконец, в пределах прозрачной перегородки лежит ядро перегородки (nucleus septi pellucidi) (см. рис. 37, 21).
Кроме перечисленных базальных ядер в лимбическую систему входят: кора поясной извилины лимбической доли больших полушарий, гиппокамп, мамиллярные ядра гипоталамуса, передние ядра таламуса, структуры обонятельного мозга.
Дата добавления: 2016-01-29; просмотров: 1655;