Экспериментальные эволюционные машины (ЭЭМ)
В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.
Суть непрерывного процесса заключается в постоянной подаче питания в зону развития популяции и одновременном отборе избыточной части из рабочего пространства. Природная популяция, таким образом, осуществляя непрерывный обмен веществом с внешней средой и получая энергию извне, развивается в открытой системе, что является главной чертой ее динамики. В лабораторных условиях такой обмен осуществляется с помощью непрерывного потока питающей среды в рабочий объем ферментера и соответствующего оттока культуральной жидкости.
В классических периодических процессах нелимитированный рост популяции отмечается лишь во время экспоненциальной фазы, которая сменяется фазой замедления, т. е. торможения роста. При непрерывном культивировании можно застабилизировать рост популяции в любой точке на восходящей ветви S‑образного роста популяции, в том числе и в экспоненциальной фазе. Для этого, как уже говорилось, необходимо непрерывно подавать свежую питательную среду для популяции и удалять избыточную часть популяции из рабочего объема. Для поддержания плотности популяции в заданной точке фазы нелимитированного роста применяются различные способы управления скоростью протока. Основное их свойство – наличие обратной связи между приростом концентрации биомассы и удалением части популяции из ферментера. Эти величины должны быть равными и это равенство, а с ним и концентрация биомассы – поддерживаются с помощью автоматических измеряющих и следящих устройств. Среди таких способов первым был турбидостат, в котором концентрация клеток поддерживается на определенном уровне за счет регулирования оптической плотности культуры. Применение его ограничено работой с оптически однородными средами.
Для процессов культивирования, в которых имеется прямая связь между приростом биомассы и изменением рН‑культуры (например, при потреблении физиологически кислого источника азота), разработан и используется рН‑статный способ управления скоростью протока. При этом скорость протока с помощью автоматических устройств уравнивается со скоростью изменения рН растущей популяцией, а следовательно, и со скоростью роста, что обеспечивает поддержание концентрации биомассы на заданном уровне.
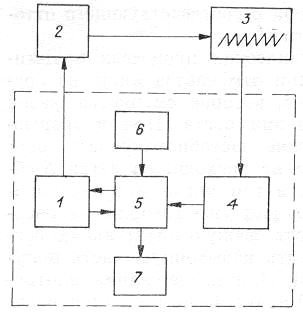
Рис. 11. Блок‑схема процессов непрерывного роста микробных популяций.
1 – культиватор; 2 – датчик‑измеритель; 3 – самописец с регулятором; 4 – система управления насосами‑дозаторами; 5 – система дозирования; 6 – сосуд с питательной средой; 7 – сборник урожая [Печуркин, 1982]. (Штриховой линией отмечена блок‑схема хемостата.)
Наиболее просто и надежно система поддержания концентрации биомассы (клеток) и управления скоростью протока работает в импульсном режиме (рис. 11). Основу схемы составляет датчик‑измеритель. Для турбидостата это – датчик оптической плотности, для рН‑стата – электродная пара рН. Измеренное значение параметра в виде электрического сигнала подается на схему управления протоком (блоки 3 и 4). Если величина параметра достигает заданной, то в системе управления вырабатывается управляющий сигнал, который передается в систему дозирования. Происходит долив порции свежей питательной среды и одновременно такой же по объему слив части суспензии из ферментера. Разбавление культуры свежей питательной средой приводит к снижению величины управляющего параметра, и проток выключается. Продолжающийся рост популяции снова вызывает возрастание величины управляющего параметра до уровня срабатывания следящей системы и т. д. На самописце 3 вырисовывается «пила», частота и размер зубцов которой определяются особенностями схемы управления и скоростью роста популяции.
В отличие от описанного выше турбидостатного культивирования в нелимитированных условиях, когда для поддержания устойчивого состояния требуются регуляторы, хемостат характеризуется обязательным внешним ограничением роста. Такое ограничение является устойчивым регулятором стационарности параметров среды и популяции. Как правило, это – лимитирование недостатком одного из компонентов питания или ингибирование роста продуктами метаболизма. Основа хемостата – подача питательной среды с постоянной скоростью протока. Популяция, утилизируя субстрат, «загоняет» себя в условия лимитирования по одному из компонентов среды, потребляя его до низких остаточных значений. Хемостат получил широкое распространение, так как простота аппаратурного оформления сочетается в нем с широкими экспериментальными возможностями.
Для нас здесь необходимо подчеркнуть два важных свойства турбидо‑ и хемостата. Первый соответствует росту популяций в нелимитированных условиях, что в природе может встречаться на ранних фазах экологической сукцессии, например при заселении новой экологической ниши. Второй – с глубоким лимитированием роста – является аналогом большинства природных ситуаций, где повсеместно встречается ограничение роста.
С точки зрения функционирования открытых систем хемостат и турбидостат – это термодинамические системы, способные находиться в устойчивых стационарных состояниях. Причем хемостат соответствует случаю постоянных потоков, а турбидостат – случаю постоянной организации (или постоянных реакционных сил). Других условий стационарности в открытых системах просто не бывает. Таким образом, в руках экспериментаторов имеются открытые системы двух основных типов развития (и для экологии, и для термодинамики). Если в таких системах будут происходить эволюционные изменения, переход от одного стационарного состояния к другому в результате изменения качественных свойств систем (например, в результате процессов мутирования и отбора), то главные характеристики этих сукцессионных перестроек, или шагов эволюции, можно измерить, не теряя общности подхода с точки зрения как биологии, так и физики. Естественно, что основу такого единства составляет энергетический подход.
Накоплено достаточно много данных по работе «эволюционных машин» обоих типов. Кинетика накопления активных мутантов, вытесняющих исходную форму из ферментера в результате процессов автоселекции, выглядит очень разнообразной (см. [Печуркин, 1978, 1982]). Здесь мы только перечислим наиболее интересные варианты.
В турбидостате это – мутанты с повышенной максимальной удельной скоростью роста; более «резистентные» мутанты, т. е. способные расти с повышенной скоростью в условиях ингибирования роста. Для хемостата характерны мутанты с повышенным сродством к лимитирующему субстрату; более экономичные формы; более жизнеспособные мутанты, т. е. с пониженной скоростью отмирания, и т. д.
В турбидостате микроэволюционный переход сопровождается увеличением скорости протока, а следовательно, и Н пад (в соответствии с рис. 10, б); в хемостате процесс автоселекции приводит к более полному использованию лимитирующего субстрата, т. е. к снижению Н неисп (в соответствии с рис. 10, в). И для обоих случаев характерно увеличение потока использованной энергии, т. е. Н исп. Таким образом, энергетические принципы однозначно указывают направление эволюционных перестроек, несмотря на кажущееся внешнее разнообразие эволюционных переходов.
Оказалось, что в хемостате и турбидостате можно хорошо экспериментировать с более сложными взаимодействиями, например изучать сосуществование видов, а также трофические отношения типа хищник – жертва, паразит – хозяин. При длительном развитии таких систем в проточных условиях наблюдалась хорошо выраженная тенденция к стабилизации их развития и коэволюции, причем Н исп возрастал.
Эволюционные эксперименты по типу турбидостата были проведены в лаборатории доктора С. Спигелмана (США) не с живыми клетками, а с макромолекулярными системами, способными к воспроизведению. Действию отбора в ряду последовательных разведений (переносов в пробирки с приготовленной для размножения средой) подвергался носитель наследственности – молекула РНК бактериофага Qβ или ее укороченные варианты. Репликация РНК осуществлялась с помощью фермента РНК‑полимеразы, выделенной из зараженной фагом культуры кишечной палочки.
Остановимся подробнее на одном из экспериментов по накоплению и идентификации мутантов, резистентных к этидийбромиду. В качестве исходной формы в опыт бралась малая реплицирующаяся молекула РНК (MDV), состоящая из 218 нуклеотидов, расположенных в известной последовательности. Реакционная смесь в каждой из 25 пробирок содержала 15 мкмоль ингибитора – этидийбромида, который способен обратимо снижать скорость удлинения цепи молекулы. В каждой пробирке, начиная с первой, реакция инкубировалась в течение 10 мин, а для переноса в последующую пробирку использовалась малая доля (10‑5) материала предыдущей. В данных условиях эксперимента происходило экспоненциальное размножение РНК в каждой пробирке, так что действие отбора было направлено на увеличение скорости репликации, которая и возросла почти в 2 раза.
Изучение последовательности нуклеотидов в мутантной РНК показало, что три нуклеотида отличаются от нуклеотидов «дикого» (исходного) типа. Каждая из трех единичных мутаций происходила в разное время. Химическая основа резистентности, по‑видимому, заключается в элиминации сайтов, связывающих этидий, благодаря специфическим изменениям в последовательности нуклеотидов. Это выражалось в том, что мутантная РНК связывала меньше этидия, чем молекулы «дикого» типа.
Эти эксперименты стимулировали теоретические исследования уже упоминавшегося нами М. Эйгена [1973] по эволюции самовоспроизводящихся структур типа гиперциклов замкнутых цепочек из ферментов и нуклеиновых кислот. Он рассматривал конкуренцию таких единиц в открытых системах при двух типах селекционных ограничений: постоянные потоки (аналог хемостата) и постоянная организация (аналог турбидостата). Близость его результатов к расчетам по кинетике эволюционных переходов в микробных популяциях оказалась удивительной. Если же дополнить его расчеты, введя поток богатых энергией мономеров, которые служат источником энергии и вещества для синтеза исследуемых полимеров, то действие энергетического принципа экстенсивного развития становится совершенно очевидным для обоих случаев селекционных ограничений [Печуркин, 1982]. Таким образом, и на неживых системах с автокатализом и внешним ограничением «работа» энергетических принципов может быть достаточно легко продемонстрирована.
К сожалению, получить данные со строгими измерениями кинетики и эволюции сложных экологических систем гораздо труднее. Трудно замерять и динамику потоков энергии. Однако некоторые результаты экологических сукцессий и эволюционных перестроек достаточно хорошо иллюстрируют действие энергетических принципов.
Мы уже говорили, что энергетические принципы и биогеохимические принципы Вернадского очень хорошо соответствуют друг другу. «Всюдность» жизни определяется «всюдностью» потока энергии, и можно только удивляться подгонке локальных круговоротов вещества к захвату солнечной энергии. В этом разделе мы рассмотрим ряд примеров, иллюстрирующих действие энергетических принципов в трех основных звеньях круговорота: автотрофах‑продуцентах, гетеротрофах‑консументах и разлагателях‑редуцентах.
Общеизвестно, что при изменении (даже очень резком) условий среды может произойти практически полная смена видов, особенно доминирующих в экосистеме, но функция круговорота страдает менее всего. Например, при сукцессионных перестройках в экосистемах работает правило, сформулированное А. Лоткой еще в 1925 г.: максимизируется энергия в биологических системах. Сукцессия сопровождается удлинением жизненных циклов, ростом степени замкнутости круговоротов вещества и в конечном счете переходом к стабилизированному состоянию, где весь прирост биомассы в автотрофном звене компенсируется ее расходом в последующих звеньях. Такой «гомеостаз» системы может быть ярко выражен. Около 40 лет назад было отмечено, что содержание хлорофилла на 1 м2 в разных сообществах бывает примерно одинаковым [Гесснер, 1949; цит. по: Одум, 1975]. То есть в целых сообществах содержание зеленого пигмента гораздо более равномерно распределено, чем в отдельных растениях или их частях. В сложившихся сообществах самые разнообразные растения – старые, молодые, освещенные, затененные, многолетние, однолетние – представляют собой единое целое и по‑своему приспособлены к захвату поступающей солнечной энергии. Растения, адаптированные к слабому свету, содержат меньше хлорофилла на единицу действующей поверхности, чем растения верхнего яруса. Степень поглощения светового потока хорошо определяется невооруженным глазом в глухом таежном лесу, в старой дубраве и в многоярусных джунглях: даже в яркий день там царит полумрак.
Содержание хлорофилла иногда предлагается считать за одну из основных продукционных характеристик природных экосистем (точнее ассимиляционное число, т. е. продуктивность на 1 г хлорофилла, выраженная в граммах кислорода за час). И для водных, и для наземных экосистем этот показатель является одним из наиболее устойчивых.
Может быть, еще ярче проявляется эта важнейшая особенность биосистем – функциональное сходство при различиях видового состава – при изучении суммарной биологической активности почв. Несмотря на явные различия в составе организмов, результирующая активность определенного горизонта, слоя или даже типа почвы остается примерно одинаковой. То же можно отнести и к дыханию, т. е. выделению углекислоты. И все это имеет место, несмотря на сложную пространственную микроочаговость почвенных ценозов. Такой микроочаг (0,05–5 мм в диаметре) иногда включает и автотрофный компонент; тогда он служит реальной природной структурой, соответствующей представлению о простейшей микробной экосистеме. Таким образом, в природной обстановке уже в пределах 3–5 мм складываются сложные сообщества, осуществляющие круговорот! И такие сообщества относительно устойчивы по функционированию.
Крупные экосистемы воды и суши с точки зрения структурной организации и наличия имеющихся видов трудно объединить по сходству. Возможно, что ни одного вида, общего для таких экосистем, и не удастся отыскать. Но с точки зрения функционирования, сукцессионной динамики и эволюции они во многом схожи. По крайней мере эволюция в пределах разных групп экосистем может количественно характеризоваться ростом эффективности использования солнечной энергии.
Из‑за низкой плотности и рассеянности потока квантов солнечного излучения даже в сбалансированных ценозах используется на фотосинтез около 1% падающего потока, а в целом по биосфере – ниже 0,5%. Это кажется очень малым, но мы теперь уже знаем, как непросто живым экосистемам в условиях нехватки вещества производить его всевозможные циклы. Мы уже описывали одно из удивительнейших чудес природы – растение, которое способно на огромную высоту навстречу солнечному лучу поднимать растворы необходимых солей, чтобы добыть энергию себе и последующим звеньям, поставляющим эти соли растению в круговороте.
Зато с каждым последующим звеном коэффициент использования энергии повышается, травоядные животные используют 10–15% от их кормовой базы, а хищники – до 30%. Поскольку консументы берут около 10% энергии растений, то, как отмечает С. С. Шварц [1980], часто делается вывод о том, что не энергетические ресурсы лимитируют развитие. Однако более тонкие наблюдения говорят, что такие выводы надо использовать с осторожностью. Оказывается, энергетический баланс животных очень напряжен. Поддержание энергетического баланса может рассматриваться как основа адаптаций.
Однако нельзя отводить растениям слишком пассивную роль. Живая природа более изобретательна, чем мы склонны считать. Одним из ярких примеров этому может служить увеличение захвата энергии растениями в эволюции. Если первичный бактериальный фотосинтез был связан с энергетически низкой длинноволновой частью спектра солнечного излучения, то современные зеленые растения используют наибольший возможный поток солнечной энергии. Считающийся более древним бактериохлорофилл имеет максимум поглощения между 800 и 900 нм, где поток солнечной энергии существенно меньше.
Говоря о действии ЭПЭР в биосфере, обратимся к выводам теоретиков. В обстоятельном труде «Самоорганизация в неравновесных системах» Г. Николис и И. Пригожин [М., 1979] затрагивают аспекты эволюции экосистем. Рассматривая вопросы устойчивости системы против структурной флуктуации с новой функцией (что‑то типа активного мутанта в популяции), авторы приходят к выводу, что «в качестве движущей силы эволюции следует рассматривать энергетическую диссипацию» и что «процессы эволюции приводят к усилению эксплуатации окружающей среды» (с. 456). Обсуждавшийся нами энергетический принцип достаточно полно и точно соответствует этим выводам.
Дата добавления: 2016-01-26; просмотров: 812;