ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ РАСТЕНИЯ
Покрытосеменные – наиболее высокоорганизованные растения на земном шаре. Наиболее характерно для них наличие цветка, в завязи которого укрыты семяпочки. После оплодотворения из завязи развивается плод с семенами внутри. Способность к видообразованию у покрытосеменных очень хорошая. Поэтому среди покрытосеменных много видов водных растений, чрезвычайно интересных по своим биологическим особенностям.
Общие экологические признаки различных видов водных растений появляются в результате естественного отбора. Основные средства приспособленности растений к современным условиям жизни в воде: 1) рассечение листовых пластинок на мелкие нитевидные доли, что обеспечивает растению увеличение поверхности при поглощении световых лучей, кислорода, углекислого газа, минеральных солей; 2) сильное развитие аэрокамер в связи с недостаточным количеством воздуха в воде, что уменьшает также плотность растения, способствующую вертикальному положению в воде и вынесению на поверхность воды листьев и цветков; 3) отсутствие устьиц за исключением верхней поверхности плавающих листьев, так как газообмен происходит через всю поверхность водных растений; 4) слабое развитие кутикулы, утратившей свою защитную функцию; 5) слабое развитие корневой системы или отсутствие ее, так как функцию поглощения воды и растворенных в ней солей выполняет вся поверхность растения; 6) отсутствие сосудов в связи с утратой необходимости проводить воду и минеральные соли, что связано с поглощением их любой частью растения (характерно только для полностью погруженных растений); 7) частичная редукция механической ткани в связи с большей, чем воздушная среда, плотностью воды; механическая ткань сохраняется только в центральных частях стебля в виде тяжей, которые позволяют растению испытывать натяжение, противостоять движению воды; 8) частичное или полное отсутствие дифференциации ткани листа на губчатую и столбчатую, что связано с недостаточным проникновением света даже на небольшую глубину; 9) сравнительно быстрый рост водных растений, благоприятные условия для которого обеспечиваются водной средой; 10) способность к вегетативному размножению намного большая, чем у наземных видов, что позволяет водным растениям выживать в сложных условиях обитания в водной среде; 11) выделение поверхностью слизи, что уменьшает трение органов друг о друга, предохраняет ткани от выщелачивания, защищает зимующие почки от охлаждения и спасает на некоторое время растения от высыхания при • понижении уровня воды. Все эти общие черты приспособленности в той или иной степени можно наблюдать у аквариумных цветковых растений.
Вместе с тем водные растения отличаются друг от друга и видовыми приспособительными признаками. Например, некоторые виды рдестов могут существовать только при связи с тремя средами – водой, почвой и атмосферой. Роголистник живет без почвы, при соприкосновении с атмосферой погибает, и ему достаточно только водной среды.
По отношению к почве, воде и атмосфере водные растения можно разделить на несколько экологических групп: 1. Жизнедеятельность происходит в основном в толще воды. 2. Необходимые условия жизни вида – вода и атмосфера. 3. Растения связаны только с водой и почвой. 4. Растения нуждаются в почве, воде и атмосфере (укоренены на дне, листья плавающие). 5. Растения частично возвышаются над водой, будучи укорененными в почве.

Рис. 6. Элодея канадская
К первой группе растений относятся элодея канадская, элодея зубчатая, роголистник темно‑зеленый, пузырчатка обыкновенная, альдрованда пузырчатая, ряска трехдольная.
Элодея канадская принадлежит к семейству водокрасовых (рис. 6). Родина – пресные водоемы Америки. В XIX в. этот вид спонтанно акклиматизировался в Англии, а затем в Европе.
Элодея канадская быстро освоила водоемы нового ареала, вытеснив многие аборигенные растения. За быстрое распространение этот вид получил название «водяная чума». В Европу попали только женские экземпляры растения (оно двудомное), позднейшие попытки целенаправленно акклиматизировать мужские экземпляры эффектов не дали, так как пригодные для обитания экологические ниши уже были заняты женскими растениями. Поэтому быстрое распространение элодеи обеспечивалось исключительно вегетативным размножением. В настоящее время элодея канадская широко распространена в водоемах европейской части СССР, а в последнее десятилетие все чаще встречается и в Сибири, например в Ангаре, в районе Иркутска.
Листья элодеи собраны в мутовки по 3 – 4, мелкие, обычно длиной 7 – 12 мм, продолговатые, по краям мелкозубчатые (это заметно только в лупу). Стебель тонкий, ломкий, длиной 40 – 60 см, ближе к поверхности воды ветвится. Цветки образуются в пазухах верхних листьев на тонкой и длинной трубке, похожей на цветоножку: у них три белых лепестка. Корневая система развита слабо, ассимиляция идет всей поверхностью растения. В природе встречается в прудах, тихих заводях речек и озер, предпочитает слабопроточные воды. В некоторых водоемах (Московская область) образует большую сырую биомассу–более 4 кг на 1 м2.
Заросли элодеи населены огромным количеством беспозвоночных пресных вод и охотно посещаются рыбами. Эти же заросли служат субстратом для икры рыб, среди ветвей элодеи первое время прячутся и кормятся стаи мальков. Растения употребляются в пищу многочисленными водными обитателями, в том числе карповыми рыбами, окунем, ондатрой. Но есть интересные наблюдения, в результате которых установлено, что прудовик обыкновенный использует в пищу 40 видов высших растений, а элодею канадскую отвергает даже при сильном голоде.
Пищевая ценность элодеи очень высока: белков она содержит более 18 %, жира – около 3 %. Каротин, содержащийся в элодее, превращается в организме животных в витамин А. Элодею используют и на корм скоту с различными кормовыми добавками. Заготавливается высушиванием, силосованием; из нее приготовляют витаминную муку. При избытке растений или при расчистке водоемов употребляется как зеленое удобрение.
При сборе элодеи на экскурсии следует обратить внимание на серый налет, обычно покрывающий листья и стебли растений. Чем выше жесткость воды природного водоема, тем больше на листьях налета. Это осадок соли кальция СаСО3, выделенный растением в процессе ассимиляционной деятельности. Световой период фотосинтетического процесса связан с поглощением растворенного в воде углекислого газа. Когда его в воде недостаточно, растения начинают вырабатывать углекислый газ из гидрокарбоната кальция:
Са(НСО3)2 = СО2
потребляется
растениями
+ H2O + CaCO3
выпадает
в осадок
Ночью, когда растения не поглощают углекислый газ, он снова накапливается в водоеме в результате дыхания растений и животных, и при взаимодействии углекислого газа с нерастворимым в воде карбонатом кальция (CaCO3) образуется угольная кислота (H2CO3). В кислой среде карбонат кальция превращается в гидрокарбонат Са(НСО3)2, т. е. реакция идет в обратном направлении. Заросли элодеи могут вызвать ночью замор – удушье рыб в результате бурного обратного процесса реакции к появления избытка углекислоты в воде.
Осенью элодея образует зимующие почки, которые опускаются на дно и пережидают зиму в состоянии покоя. Иногда внезапные холода осенью или на мелководье вызывают вмерзание неопавших почек и стеблей в лед. Куски льда с вмерзшими частицами элодеи следует поместить в холодную воду и оттаивать в прохладном помещении (быстрое оттаивание в теплой воде вызовет гибель растения). Затем оттаявшие стебли и почки элодеи выставляют на солнечное окно или освещают электролампами. Вскоре почки трогаются в рост. Перезимовавшие в природе веточки элодеи хорошо и быстро разрастаются в аквариумах. При многолетнем содержании в аквариуме элодея постепенно мельчает, замедляет рост. Для содержания в школьном аквариуме следует рекомендовать более красивый и легко разрастающийся в искусственных водоемах вид элодеи из Аргентины – элодею зубчатую*, или эгерию. Зубчатая элодея – тропическое растение, не имеет сезонных циклов и растет в аквариумах круглый год, легко размножается вегетативно. Стебли длинные, более толстые, ярко‑зеленые. Заостренные на концах ланцетовидные листья собраны в мутовки, которые расположены тем чаще на стебле, чем ярче освещение. Длина листьев до 15 мм, стебли достигают в длину 2 м. Хорошо растет при температуре воды от 16 до 30°С, у поверхности воды стебли обильно ветвятся. Растение очень ломкое, большое количество обломков стеблей выделяет в воду сок, который содержит, по‑видимому, ядовитые вещества (улитки это растение не едят). Элодея зубчатая может свободно располагаться в толще воды и при этом хорошо себя чувствовать, но аквариум выглядит красивее, если элодея посажена пучками стеблей. При посадке нужно прижать их нижние концы камнем.
* Элодея зубчатая выделена в самостоятельный род эгерия.
Корни у элодеи немногочисленные, в виде отдельных белых неветвящихся нитей, функция которых – закрепление растения на субстрате. Функцию поглощения выполняет вся поверхность тела.
Элодея используется в курсе ботаники и общей биологии для рассмотрения пластид при большом увеличении микроскопа. Это округло‑овальной формы образования зеленого цвета. В теме «Лист» используют элодею для демонстрации мутовчатого расположения листьев, тем более что среди комнатных растений, используемых при изучении листорасположения, очень редко встречаются растения с мутовчатым расположением листьев. При изучении вегетативного размножения растений на уроке можно использовать результаты опыта, поставленного в уголке живой природы.
С этой целью за месяц до урока надо отрезать и поместить в хорошо освещенный сосуд верхушки стебля, черенок стебля снизу, часть стебля с двумя мутовками, часть стебля с одной мутовкой, отрезок стебля между мутовками. За месяц от этих отрезков (кроме последнего) начинают расти новые стебельки.
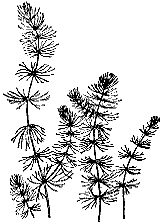
Рис. 7. Роголистник темно‑зеленый
Роголистник темно‑зеленый (семейство роголистные) – одно из самых распространенных растений в пресных водоемах нашей страны и представляет исключительный интерес как пример полной адаптации к водной среде (рис. 7). Роголистник – единственное из наших водных растений, цветущее под водой. Как и другие растения этой группы, роголистник утратил тесную связь с почвой, корней у него нет. Изредка можно наблюдать отходящие от стеблей белые неветвящиеся нити, но они представляют собой видоизмененные ветки, играющие роль якоря.
Листья роголистника темно‑зеленые, вильчато рассечены на 2 – 4 доли. На одной из сторон мутовками расположены беловатые зубчики. При демонстрации растения следует обратить внимание на радиальную симметричность мутовок. Стебли тонкие, ломкие, особенно к концу лета, тянутся на 30 – 80 см, густо ветвятся. Роголистник образует подводные леса в озерах, лесных болотах, речных заводях. Вся масса зарослей легкоподвижна, поэтому на быстрых течениях не растет, растения уносятся водой. Изредка летом при быстром реете отдельные верхушки игольчатых частей листа пронзают поверхностную пленку воды и выходят в воздушную среду, но остаются живыми, пока не обсохнут. Это единственная и не обязательная связь растений с воздушной средой. Цветет на широте Москвы в июле – августе. Цветки однодомные, мелкие, невзрачные, сидят в пазухах листьев. Тычиночные цветки образованы зелеными листочками околоцветника и тычинками (10 – 12, иногда больше), пестичные имеют 8 – 12 листиков в околоцветнике и один пестик. Пыльники на 1/3 заполнены воздухом. Созревая, они отрываются и благодаря воздухоносной полости всплывают (сравните с валлиснерией), но не до поверхности, а держатся около нее. В момент подъема пыльник раскрывается, пыльца попадает в воду. Ее плотность такая же, как у воды, поэтому она остается в стоячей воде на том уровне, где вышла из пыльника. Таким образом, поднимающиеся к поверхности пыльники заполняют пыльцой всю толщу воды до поверхности, а слабые течения в воде, волны в глубине от проплывших рыб и насекомых перемещают пыльцу в слоях воды горизонтально. На рыльцах пестичных цветков есть клейкие вещества, нерастворимые в воде, к ним прилипает пыльца. Опыление, следовательно, происходит легко, и с августа зреют снабженные шипиками плоды.
Роголистник легко размножается вегетативно. От каждого кусочка стебля с мутовкой листьев могут начать жизнь новые растения. Эти обломки легко переносятся птицами и другими животными, а также лодками из водоема в водоем. Зимует растение подо льдом, к осени вся заросль утяжеляется и опускается на дно. Верхушки стеблей, тесно сжимая верхушечные листья, образуют своеобразную почку, которая может зимовать на стебле, а может отвалиться и опуститься на дно. Все эти особенности роголистника способствуют очень раннему пробуждению растения весной по сравнению с другими гигрофитами и быстрому заселению новых территорий и водоемов. Поэтому в ряде мест роголистник – главное растение водоема, затеняет и подавляет другие виды.
Роголистник образует летом большую биомассу: в районе Москвы в сырой массе – 4 кг/м2, а в озере Севан – до 20 кг/м2. В Средней Азии биомасса роголистника достигает 300 т/га. В природе служит субстратом для многих водных животных. В его зарослях скрываются мальки промысловых рыб, промысловые грызуны
используют это растение в качестве корма. Измельченный и смешанный с мучной пылью роголистник, используется в корм домашней водоплавающей птице. В зарослях роголистника обильно развиваются личинки комара. В связи с ликвидацией в СССР очагов малярии такая особенность этого растения для нас не особенно важна, но в других странах, где малярия еще имеется, например в Индии, скопления больших зарослей роголистника создают благоприятные условия для развития личинок малярийного комара, обеспечивая им защиту и не препятствуя разрастанию фитопланктона, служащего им пищей.
Для аквариума из природного водоема роголистник лучше брать летом и не целые побеги, а молодые, хорошо развитые их верхние части. Период акклиматизации обычно длится до осени, но, пережив его, растения могут хорошо перенести зиму, хотя листья мельчают и все растение утрачивает свой прежний привлекательный вид. Во избежание гибели растений рекомендуется по мере их роста, отрезать и удалять из аквариума нижние части ветвей. Взятые из природы осенью побеги роголистника, как правило, не доживают до весны.
Пузырчатка обыкновенная (семейство пузырчатковые) распространена в европейской части СССР, Сибири, Средней Азии, на Кавказе и Дальнем Востоке. Растет в канавах, прудах и других водоемах с медленно текущей водой. Длинный ветвистый стебель без корней свободно плавает в воде около поверхности и бывает иногда длиной 1 м. Листья рассечены на нитевидные доли, некоторые из них снабжены пузырьками – видоизмененными долями листа. Пузырьки, сидящие на ножках, бледно‑зеленого цвета, сжаты с боков, боковые стенки полупрозрачны. На пузырьке имеется отверстие, усаженное по краям жесткими, острыми щетинками, нижний край отверстия утолщен и имеет плотный выступ, вдающийся внутрь пузыря, с верхнего края отходит тонкий прозрачный клапан, который, смыкаясь с выступом нижнего края, плотно закрывает отверстие. Клапан при малейшем давлении извне вдается внутрь и открывает отверстие, но вследствие упругости при прекращении давления сразу возвращается на старое место, перекрывая вход. Изнутри открыть клапан невозможно из‑за выступа нижнего края отверстия. Эти пузырьки – ловчие камеры. Натыкаясь на клапаны, мелкие водные животные открывают клапан, проникают в пузырь и оказываются в западне (рис. 8).
Пузырчатка использует животные белки, так как воды, в которых она обитает, отличаются бедностью азотистых соединений. Однако причислить пузырчатку к растениям‑хищникам можно лишь с некоторой оговоркой. «Охота» пузырчатки пассивна. Животное само нажимает на клапан и оказывается в пузырьке, выход из которого невозможен из‑за конструкции клапана. Животное задыхается, погибает и разлагается. Образующиеся при разложении соки поглощаются стенками пузырька. В этом смысле пузырчатка существенно отличается от настоящих растений‑хищников, например росянки, что дало повод Ч. Дарвину метко окрестить пузырчатку растением‑стервятннком (питающимся падалью). Стенки пузырька выделяют особые ферменты и бензойную кислоту, предохраняющие погибшую жертву от загнивания, вызываемого микроорганизмами. Таким образом пузырчатка как бы консервирует жертву, обеспечивая длительное ее усвоение.

Рис. 8. Пузырчатка обыкновенная
Цветет пузырчатка летом. Желтые цветки поднимаются на высоких тонких цветоножках над поверхностью воды. Они неправильной формы, двугубые со шпорцем. Нижняя губа венчика длиннее верхней. Осенью листья на концах стеблей прекращают рост и складываются шаровидно, образуя простейшую зимующую почку. Растение опускается с холодами на дно, теряет из полостей воздух, но почки сохраняют воздушные полости. Поэтому почка при зимовке направлена вверх. Весной она отрывается от гниющего стебля, всплывает и быстро начинает расти в теплых поверхностных слоях воды на ярком освещении.
В аквариумах пузырчатка хорошо живет в летние месяцы и при ярком солнечном освещении иногда зацветает. Сохранить в аквариуме пузырчатку обыкновенную зимой трудно. Но если остались почки, взятые осенью в природном водоеме, их держат в сосуде с водой на окне при температуре на 2 – 3 °С ниже комнатной. С наступлением весны при дополнительной подсветке почки трогаются в рост и молодые растения переносятся в аквариум. Учащиеся могут наблюдать пузырчатку обыкновенную в аквариуме от почки до образования новых почек.
Большинство растений семейства пузырчатковых распространены в тропических пресных водоемах, из них 2 – 3 вида прочно акклиматизировались в любительских аквариумах. Чаще других в аквариуме содержат пузырчатку горбатую. От пузырчатки обыкновенной она отличается более тонким и гибким стеблем и очень узкими и короткими листочками. Зимой хорошо чувствует себя в аквариуме при достаточной подсветке и температуре около 20 °С.
В пресных водах СССР обитает настоящее растение‑хищник альдрованда пузырчатая. Это родственница широко известной росянки круглолистной, обитающей на болотах. Оба растения принадлежат к семейству росянковых. Альдрованда пузырчатая в СССР встречается на Украине, Кавказе, в дельте Волги, местами в Курской, Ленинградской областях, а также в Средней Азии. Растение плавает у поверхности, корней нет, листья на горизонтальном стебле расположены по 8 – 9 мутовками, черешок клиновидный, плосковатый, на конце ложковидная пластинка, которая может складываться. Возле пластинки длинные чешуйки. Обычно створки пластинки‑клапана составляют угол 60°. Край покрыт острыми щетинками, затем идет зона выделяющих слизь железок, зона без железок и, наконец, зона с переваривающими железками и чувствительными щетинками. При складывании пластинок две зоны плотно смыкаются, а вогнутая – образует полость.
Механизм активной охоты у этого растения таков. Щетинки воспринимают раздражение и приводят в движение аппарат сведения пластинок. При раздражении только внешних щетинок пластинки сходятся медленнее, при раздражении и внутренних – быстрее и плотнее. Створки замыкаются герметически. Пойманное мелкое животное бьется в полости, своим движением раздражая щетинки и вызывая еще более сильное смыкание створок. Это же раздражение запускает в действие пищеварительные щетинки, они выпускают особый фермент, убивающий и переваривающий добычу. По завершении поглощения соков возбуждение спадает и створки раскрываются, непереваренные остатки смываются с пластинок водой.
Альдрованда хорошо растет и вегетативно размножается в аквариумах с верхним освещением лампами накаливания. Скопления пузырчатки у нас и в тропических водоемах– благоприятный биотоп для развития личинок комаров. Ареал альдрованды разорван не только на территории СССР, но и за ее пределами (местами в ГДР, Польше, Северной Италии, Южной Франции, в болотах Бенгалии, в тропической части Восточной Австралии). Это растение находится под охраной.
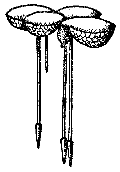
Рис. 9. Ряска трехдольная
Из семейства рясковых на территории СССР обитает пять видов. Ряска трехдольная живет целиком погруженная в воду. Ряска трехдольная в условиях школы – хороший объект наблюдения (рис. 9). Интересны ее морфология, оригинальный способ вегетативного размножения, условия перезимовки в аквариуме, значение для других обитателей искусственного водоема. Остальные виды рясок живут на поверхности водоема, соприкасаясь, как с водой, так и с атмосферой: ряска малая размером всего 2 – 3 мм, ряска горбатая 5 – 6 мм, спироделла многокорневая и вольфия бескорневая – 1,5 мм.
Зеленые пластинки рясок – это не листья, а видоизмененные стебли, принявшие на себя функцию фотосинтеза. Стебли ряски сравнивают с листьями кувшинок или водокраса: интересна конвергенция различных органов растений разных семейств, находящихся в сходных условиях обитания.
На экскурсии и на занятиях кружка следует внимательно рассмотреть растение. Стебель безлистный, округлый (у ряски малой – овальный). Из центра пластинки снизу отходит один полупрозрачный корешок (у многокоренной – их несколько). Конец корешка заключен в конусовидный кармашек – противовес. Корешки обеспечивают устойчивость растения на поверхности водоема. Под лупой в тканях пластинки видны аэрокамеры, наполненные воздухом. Они способствуют плавучести пластинки. Проверить это положение можно на занятии – осторожно пинцетом взять ряску за корешок и заглубить. Отпущенная пластинка ряски всплывает. Летом пластинки ряски многокоренной снизу красные.
Красная окраска многих гигрофитов объясняется присутствием в клетках пигмента антоциана. При ярком освещении хлорофилл частично разрушается и антоциан экранирует хлорофилл от излишнего света. Колпачок ряски подобен по происхождению корневому чехлику наземных растений, но специфика обитания в воде привела к редукции внутреннего строения чехлика: исчезли подвижные крахмальные зерна – статолиты, осталась лишь внешняя часть – оболочка. Один из представителей рясковых – вольфия утратила и корешок.
Цветут ряски чрезвычайно редко. В связи с этим до начала XVIII в. даже ботаники относили ряски к водорослям*. Цветки ряски появляются на краю пластинки – стебля. Один женский и два мужских цветка сидят как бы в мешочке, и это соцветие рассматривают как крайне упрощенный початок. Найти цветок ряски – большая удача даже для ботаника. Он считается самым маленьким цветком в мире. За последние 200 лет в скандинавских странах зарегистрировано 33 случая цветения, в Польше – 2, в нашей стране – 26.
* Впервые обнаружил цветение ряски итальянский ботаник Валлиснери.
Плод ряски размером с маковое зернышко снабжен килем, с помощью которого недолго плавает и через один‑два дня прорастает.
Обычно ряски размножаются вегетативно. На краю пластинки образуется почка, затем из нее развивается пластинка.
Зимой ряски хорошо растут в аквариумах, освещаемых электролампами. В природе пластинки опускаются под лед. Происходит это потому, что масса ряски равна массе воды при температуре +4 °С. По мере охлаждения воды аэрокамеры наполняются водой, ряска становится все тяжелее и, опускаясь, избегает вмерзания в лед. Быстрое развитие рясок может вызвать нежелательные последствия для других обитателей водоема, так как ряски препятствуют проникновению солнечного света и кислорода сверху, отчего страдают погруженные высшие растения, фитопланктон и животные, которым для дыхания необходим атмосферный кислород.
Умеренное развитие ряски создает благоприятные условия для жизни многих водных организмов. Ряски значительно способствуют обогащению воды кислородом, что можно проверить в условиях уголка живой природы.
Ряска как в природе, так и в аквариуме служит хорошим убежищем для многих беспозвоночных животных и мальков рыб. Многие животные употребляют ряску в пищу (рыбы, птицы, ондатра и др.). Ряска используется и как дополнительный корм свиньям и домашней птице. Особой питательностью отличается ряска малая. В сухой массе ее содержится до 25 % банков, до 4–5 % жира, 30 % безазотистых экстрактивных веществ, до 20 % клетчатки и 18 % золы.
Ряску заготовляют и как добавку к силосу, используют также для приготовления кормовой муки. Употребляют ее и в качестве зеленого удобрения.
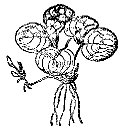
Рис. 10. Водокрас лягушачий
Вторую группу составляют водокрасы, лимнобиумы и телорезы. Водокрас, или лягушатник (семейство водокрасовые), – обычное растение стоячих и слабопроточных вод Средней России. Встречается даже в мелких водоемах и дренажных канавах вдоль шоссе (рис. 10).
Водокрас имеет округлые с сердцевидным вырезом у основания ярко‑зеленые выпуклые пластинки листьев, лежащие на поверхности воды. При ярком солнечном освещении на поверхности листьев проступают пунцовые пятна антоциана или же листья приобретают общий оливково‑коричневый оттенок. При освещении молодые листья могут быть розовыми, красноватыми – антоциан защищает в молодых листьях хлорофилл от разрушения избыточным освещением. Черешки листьев длиной 5 – 10 см отходят розеткой от короткого стебля 1 – 2 см. Вниз от стебля опускаются немногочисленные толстые неветвящиеся белые корни, покрытые белыми волосками. От стебля отходят и боковые побеги, из почек на их концах развиваются дочерние кустики. Часто, потянув за одно растение, можно вытащить целую цепь из 10 – 15 растений‑потомков. Осенью на концах столонов почки не развиваются (задержку вызывают сокращение освещенности и холодная вода), а обилие питательных веществ делает такую почку более тяжелой. Оторвавшись от материнского растения, почки плавно опускаются ко дну и останавливаются у поверхности грунта. Весной с увеличением освещенности и температуры воды активизируется фотосинтетический процесс в почках, в специальных полостях накапливается воздух, почка становится легче воды, устремляется к поверхности и вскоре выпускает первый плавающий лист. Водокрас может зимовать в аквариумах при электроосвещении, но сильно мельчает зимой. Гораздо устойчивей для аквариумной культуры его американский родственник лимнобиум, или трианея, который при электроосвещении хорошо растет круглый год.
Рассматривая водокрас или лимнобиум, обратите внимание на лист. Лист обеспечивает связь этого пришельца в воду с воздушной средой. Поверхность гладкая, выпуклая, покрыта восковым налетом, отчего лист не смачивается (показать это можно, слегка заглубив лист палочкой в воду – он тотчас всплывет на поверхность; старые листья постепенно погружаются, отслужив свою службу). На верхней стороне много устьиц, испарение идет непрерывно: во‑первых, не надо экономить влагу, как наземным растениям; во‑вторых, непрерывное испарение обязано сильному восходящему току от корней к листьям. Если чистое, с белыми корнями растение поместить в мутную воду, уже через час волоски на корнях покрываются частицами мути, что показывает, с какой силой тянут корни воду.
В аквариумах с плотным покрытием поверхности воды листьями водокраса или лимнобиума листья располагаются на поверхности воды горизонтально: молодой, свернутый лист начинает еще под водой отклоняться в сторону от вертикальной оси. Отошедший в сторону черешок выносит постепенно разворачивающийся лист к поверхности по касательной. При плотной засадке лист выходит наполовину из воды еще свернутым, черешок уплотнен, острие свернутого листа действует наподобие шила, протыкая закрытую листьями поверхность, отклонение листа в сторону от оси стебля незначительно. Сначала лист поднимается наполовину из воды, затем начинает отклоняться на черешке, а листовая пластинка одновременно разворачивается. Таким образом каждый молодой лист оказывается обязательно над более ранними листьями, обеспечивая растению прогресс в росте и завоевании пространства. Эту приспособленность водных растений с плавающими листьями к разной плотности посадки можно использовать при объяснении материала по экологии в курсе общей биологии.
Розетки лимнобиума в аквариуме редко превышают в диаметре 5 – 8 см. Тропическую красоту этого растения можно увидеть теплой весной и летом в водоеме, плотно прикрытом парниковой рамой, на ярком солнце, Тогда маленький лимнобиум достигает в диаметре 20– 25 см, а листья приобретают пунцовую окраску.
Телорез сабуровидный (семейство водокрасовые) широко распространен в пресных водоемах нашей страны и интересен прежде всего своим историческим возрастом – близкие ему формы жили еще в третичный период.
На краях листьев расположены жесткие иглы, которые могут больно поранить тело человека. По сходству с известным комнатным суккулентом алоэ (по внешнему виду) распространено и другое его русское название – водяное алоэ. Листья телореза жесткие, ломкие, саблевидно изогнутые, в сечении (особенно у основания) трехгранные, конец сильно заострен. Листья собраны в густую розетку от короткого мясистого толстого стебля (длина его до 1,5 см, диаметр до 5 см), внизу которого расположено и жесткое корневище. Корни – длинные белые нити без разветвлений, обычно 3 – 6 нитей длиной до 30 см. Общий размер растения без корней до 40 – 45 см. Размножается преимущественно вегетативно. Телорез – двудомное растение, но встретить одновременно в водоеме и тычиночные и пестиковые экземпляры довольно трудно: обычно бывают либо те, либо другие. Цветки крупные (диаметром до 2,5 см), белые, трехлепестковые, мужские со «многими тычинками, женские с пестиком, окруженным недоразвитыми тычинками (стаминодиями). Вегетативное размножение осуществляется образованием дочерних кустиков на побегах, которые появляются из влагалищных почек. Осенью на материнском кусте образуются зимующие почки, но отделяются не они, как у водокраса, а листья материнского растения. Отпадают и его корни. Остается корневище с почками, оно опускается на дно для зимовки. Весной из почек прорастают новые кусты, они используют питательные вещества корневища. В разгар лета колючие листья телореза покрывают огромные пространства водоемов. Вегетативное размножение вполне обеспечивает биологический прогресс этого вида и свидетельствует о его высокой стойкости: часто под густым покровом телореза гибнет другая водная растительность.
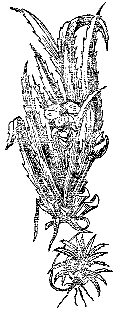
Рис. 11. Телорез алоевидный
Интересен механизм вертикальной миграции телореза.
Весной, с увеличением освещенности, возрастает фотосинтетическая деятельность в почках, и они заполняются воздухом. Зимующее корневище становится легче воды и всплывает к поверхности. Из нескольких почек на одном корневище образуются листовые розетки. Они плавают около поверхности. Листья светло‑зеленые, погруженные, ломкие. Разрастаются свободно свисающие вниз корни. Созреванию бутонов предшествует утолщение листьев, они уже не так саблевидно изогнуты, концы их направляются вверх, все большее число новых листьев появляется над поверхностью воды, сначала кончиками, а потом и на всю длину. В толстых надводных листьях увеличивается количество аэрокамер, что вызывает приподнимание всего куста за пределами водной среды (рис. 11). Цветки раскрываются над водой. После оплодотворения или отцветания всех цветков этого лета растение начинает медленно опускаться в воду. Надо обратить внимание и на такие особенности водяного алоэ: при погружении корни растения могут коснуться грунта и тогда врастают в него, препятствуя последующему всплытию; при образовании густой переплетенной дерновины в результате бурного вегетативного размножения телорезы могут и не погружаться после цветения. В обоих случаях форма куста меняется: вместо розетки раскинутых в стороны листьев они направлены вверх, и лишь слегка расходятся в стороны их концы. Такие растения могут нормально существовать либо укорененными в грунте, либо в сообществе поддерживающих друг друга растений. Отдельный такой куст плавает у поверхности на боку, а иногда и вверх корнями. Естественно, что вертикально мигрировать он не может.
К осени с уменьшением освещенности телорез вновь приподнимается к поверхности воды, но, как правило, уже не выходит листьями в воздушную среду. Летние надводные листья из‑за обильного образования антоциана могут приобретать вишнево‑свекольный оттенок и заросли телореза представляют в этот период изумительное по красоте зрелище.
Телорез интересен не только как экскурсионный объект, но и как объект работы в уголке живой природы. Из природного водоема можно перенести в школьный аквариум растения на разных этапах вегетации. Содержать их надо в чистой прозрачной воде, на ярком освещении (желательно использовать электроподсветку). К сожалению, телорез в аквариуме долго не живет, более или менее быстро погибает. Но можно сохранить растения при хороших условиях несколько месяцев. Поиски оптимальных условий наиболее длительного сохранения этого растения в аквариумах – одно из возможных заданий во внеклассной работе. Получение из собранных осенью со дна водоемов корневищ с почками новых растений ранней весной – другое задание. Вертикальной миграции телореза в аквариуме пока не наблюдалось. Возможно, что кому‑то из юннатов удастся наблюдать и зафиксировать это явление.
Телорез, водокрас и валлиснерия – три представителя семейства водокрасовых – имеют общего предка. Они в разное время и в разных условиях приспосабливались к существованию в водной среде, чем объясняется их анатомическое и экологическое различие. Как и элодея, телорез может выделять необходимые вещества из бикарбонатов, при этом листья его покрываются серой известковой корочкой. Опадая на дно летом, опускаясь с гибнущими листьями осенью, эти отложения пополняют осадочный слой так называемого озерного мела. Известковую корочку на листьях можно предложить учащимся обнаружить на экскурсиях. Измельченный и смешанный с отрубями или картофелем телорез с большим успехом применяется для откорма свиней. Летом растение быстро наращивает биомассу за счет активного вегетативного размножения, а густые плавающие дерновины этого растения можно легко собирать и заготавливать с поверхности водоемов. Урожайность телореза достигает к концу лета 10 – 13 кг биомассы на 1 м2 поверхности водоема. Зола растения содержит 30,8 % калия, 10,7 % извести. К сожалению, этот вид добавки к кормам в животноводстве пока используется очень слабо.
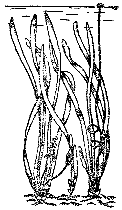
Рис. 12. Валлиснерия гигантская
Третью группу аквариумных растений представляют растения рода валлиснерия и уруть. Почва – один из необходимых факторов жизни этих водных растений в природе. В связи с этим в аквариумных условиях при их содержании нужно уделить внимание грунту. Можно в отдельных случаях растения сажать в горшочки с землей и сверху засыпать ее слоем песка.
Валлиснерия спиральная – широко известное аквариумное растение семейства водокрасовых. В культуре несколько видов. Валлиснерия встречается в водоемах тропиков и субтропиков, в СССР – в дельте Волги, на юге и юго‑западе европейской части Союза, в Средней Азии (бассейны Амударьи и Сырдарьи, в ряде озер и даже в опресненных морских заливах). Листья ярко‑зеленые, линейные, центральная жилка не заметна, конец листа закруглен, в лупу видны по кромке конца зубчики. Корневая система развита, корни тонкие, белые, узловатые, расположены пучком. Размножается боковыми побегами, на концах которых почка. Из нее развивается дочернее растение, от него через некоторое время отходит новый побег и т. д. В природе достигает в высоту 1 м при ширине листьев до 8 мм, в аквариуме мельчает (до 30 – 40 см). В аквариумах любителей распространены еще 4 вида. Один из них имеет спирально закрученные плотные листья, растет и размножается сравнительно медленно – азиатская валлиснерия. Два вида представлены гигантскими растениями – гигантской валлиснерией из Новой Гвинеи (рис. 12) и красной валлиснерией из Флориды и Кубы – валлиснерией неотропикалис. В аквариумах длина листьев у обеих достигает 2,5 м при ширине листа до 2 см. Первая имеет зеленые листья с характерным утолщением в виде валика по краю, у второй валика нет, лист коричнезо‑красный, вишневые жилки образуют на нем красивый рисунок. В аквариумах валлиснерии четырех видов растут без проблем, кроме случаев соприкасания воды с ржавчиной; замедленно растут они и в очень мягкой, кислой воде.
На примере валлиснерии можно показать приспособленность растения к жизни в водной среде. Листья лишены несущих прочных конструкций и без воды сразу опадают, быстро вянут. В то же время лист обладает большой прочностью на растяжение – течению воды, водным животным не так‑то просто его порвать. Без воды, в полупогруженном состоянии это растение не может существовать. Оно – настоящее водное. Но валлиснерия не утратила связи с воздушной средой и выносит к поверхности генеративные органы. Растение двудомное, цветки – на концах подводных цветоножек и заключены в особые пузыри из двух плотно сомкнутых прицветников. В пузыре пестичного растения – один цветок, в каждом пузыре тычиночного их несколько. Тычиночные цветки очень мелкие, и те и другие невзрачные. Женская цветоножка удлиняется и выносит цветок к поверхности, где он раскрывается в воздушной среде. Тычиночные цветки постепенно отрываются и всплывают. В воде они похожи на шарики, на поверхности цветок раскрывается. Ветер подгоняет его к женскому цветку, раскрытому чуть выше поверхности воды. Пыльца попадает на рыльце, оплодотворенный цветок уходит под воду за счет спирального скручивания цветоножки (отсюда и название – валлиснерия спиральная). Плоды развиваются у самого дна. В культуре обычны пестичные растения, за исключением красной валлиснерии (встречаются экземпляры обоих полов). Поэтому оплодотворение в аквариумах увидеть трудно, хотя цветут растения регулярно. Чаще цветение наблюдается в аквариумах, глубина которых превышает 60 см.
Валлиснерия – незаменимый объект при демонстрации на уроках выделения растением кислорода в процессе фотосинтеза. Описанный в учебнике ботаники процесс выделения кислорода элодеей на свету не всегда успешно проходит. Более удачно осуществляется этот процесс у валлиснерии. Для наблюдения надо взять сильный куст валлиснерии и срезать у него верхнюю часть листьев. Вскоре из сосудов на срезе к поверхности устремляется цепочка мелких пузырей. Это выделяется кислород.
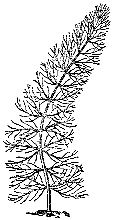
Рис. 13. Перистолистник разнолистный
Уруть, или перистолистник,– растение семейства сланоягодниковых: его листья значительно мягче по сравнению с жесткими иглами роголистника. Вынутый из воды, роголистник сохраняет свою форму – эластичные листочки урути спадают и слипаются в кисточку. Впрочем, это касается только подводных, нежных, перисторассеченных листьев. Стебли этого растения, достигнув поверхности, могут выходить в воздушную среду. Особенно это характерно для тропических видов перистолистника. В этом случае происходит сложная перестройка анатомии растения, в стеблях образуются твердые несущие тяжи, листья укорачиваются и в воздушной среде сохраняют упругость (рис. 13).
* См.: Катанская В. Высшая водная растительность континентальных водоемов. М., 1981, с. 5 – 11; Кокин К. Экология высших водных растений. М., 1982. с. 84 – 107.
На примере урути, которая иногда выносит свои стебля на поверхность воды, видно, что резкой границы у экологических групп водных растений нет*. В каждой группе можно встретить растение, имеющее некоторые черты, свойственные представителям другой экологической группы. В данном случае уруть иногда бывает связана с воздушной средой, как растения четвертой группы, о которой речь пойдет дальше. Как и большинство водных растений, уруть размножается преимущественно вегетативно, ветвлением стеблей. Отдельные веточки, кусочки стебля с одной мутовкой листьев дают начало новому растению. Количество перисторассеченных листьев в мутовке служит видовым признаком. Корневая система развита слабо, обычно это толстые белые извилистые корни‑якоря. У больших кустов с сильным разветвлением многих стеблей развивается и довольно мощная корневая система. Выход верхушек стеблей над поверхностью воды также стимулирует развитие корневой системы растения.
Осенью уруть образует зимующие почки – верхушки стеблей с плотно сложенными листьями. Эти почки могут оставаться на растении, но чаще стебель в их основании надламывается и они опускаются на дно. Собранные осенью в водоемах, эти почки быстро двигаются в аквариуме в рост и растут круглогодично, не останавливая рост и зимой, если аквариум освещается электросветом. Тропические виды перистолистников хорошо растут при электроосвещении аквариума тоже круглогодично.
Поднятые над водой стебли несут зачатки цветочного колоса. Цветки разнополые, в нижней части колоса – пестичные цветки, в верхней – тычиночные. Опыление происходит либо при помощи ветра, либо при колебании плавающих колосков волнами. Есть сведения, что порой цветки опыляются и в воде, когда колос не достигает поверхности.
Среди представителей четвертой группы аквариумных растений, связанных с водой, почвой и атмосферой, интересна кабомба. Сейчас это одно из распространенных и хорошо известных аквариумистам растений. Родина его – субтропические и тропические районы Америки. Род кабомба представлен в аквариумной культуре рядом видов, точное определение которых в условиях школьного аквариума излишне. Этот род образует самостоятельное семейство кабомбовых, а ранее был включен в семейство нимфейных.
Внешне растение состоит из тонких, длинных (до 2 м) стеблей, более или менее густо облиственных. Листья глубоко рассечены, расположены супротивно; у некоторых видов в мутовках по три листа. Около поверхности растение образует надводные белые или желтоватые небольшие цветки на коротких цветоножках. Почему же это столь своеобразное, более похожее на перистолистник растение объединялось с нимфеями? Внешний вид нимфей‑кувшинок хорошо знаком каждому школьнику. Дело в том, что у кабомб тоже есть похожие на нимфейиые плавающие листья, они невелики, возникают на верхушке стебля перед самым цветением.
Представители пятой группы – земноводные растения стрелолист и гигрофила.
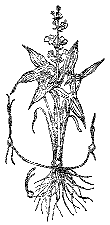
Рис. 14 Стрелолист обыкновенный
Стрелолист обыкновенный (семейство частуховые) – типичное земноводное растение нашей страны. Растет как в воде у берегов, так и в прибрежной зоне. Онтогенетические этапы развития стрелолиста соответствуют определенным сезонным изменениям. Семена прорастают весной на хорошо прогреваемом мелководье, на заливных лугах, в поймах или осенью в русле небольших речек, водоемов. Молодые, полностью погруженные листья очень похожи на листья валлиснерии – линейные, гибкие, мягкие (явление конвергенции). Отличить эти два растения в аквариуме иногда трудно с первого взгляда. У всех стрелолистов (аквариумные культуры растений этого рода называют обычно сагиттарии) более или менее четко проглядывается центральная жилка листа, конец заострен более резко, чем у листа валлиснерни. Линейные листья – это ювенильная листовая форма.
У стрелолиста обыкновенного по мере развития растений листья все удлиняются, и в русле рек доходят до поверхности, стелются около нее, иногда и на ней (верхняя часть концов листьев выходит на поверхность, высыхает и приобретает несмачиваемость). На этом этапе развития растения достигают солидной длины (например, в протоках Невы – до 1,5 м). Затем на смену мягким линейным выходят на мягких черешках более плотные, овальные, несмачиваемые сверху листья: они плавают на поверхности воды. Жесткие несущие черешки листа овальные, а позднее ложкообразные и поднимаются над водой. Последняя стадия – появление стреловидных листьев. Вслед за ними растение выносит над водой соцветие (рис. 14). Когда спадает вода, растения сбрасывают лентовидные листья и на упругих несущих черешках образуют овальные, а потом стреловидные листья. В зависимости от уровня воды это может произойти на разных этапах онтогенеза. Поэтому в русле рек стрелолист образует мощные подводные кусты высотой до 2 м с лентовидными листьями, цветки поднимаются над водой. На осушенной территории растут и большие и мелкие растения со стреловидными листьями, многие из них так и не успевают зацвести до холодов.
В аквариумной культуре несколько видов сагиттарий (стрелолистов) – стрелолист плавающий, распространенный широко по территории СССР, ряд видов из тропической и субтропической Америки. Плавающая сагиттария перед цветением – и то не всегда – образует овальные плавающие листья. Большинство же сагиттарий в аквариумах сохраняют ювенильную форму линейных листьев. Для получения надводных форм листьев надо усилить освещение и постепенно снизить уровень воды, сохраняя высокую влажность воздуха, в этом случае образуются овальные листья. В аквариуме хорошо наблюдать за перестройкой конструкции черешка и листа при переходе от погруженного положения к полупогруженному.
Стрелолист обыкновенный широко используется в хозяйственной деятельности человека. Клубни его содержат 27 – 33 % крахмала, 10 – 11 % белков, свыше 3% сахара, 0,44 % жиров. В СССР практически в пищу не употребляется (употреблялся ранее на Руси), а в Китае и Японии клубни стрелолистов готовят в печеном и вареном виде, один из подвидов стрелолиста специально культивируют. Вареные клубни по вкусу напоминают каштаны. В Китае из клубней делают муку. Высушенные, обжаренные и размолотые клубни хорошо заменяют кофе. Мучнистые корневища идут на корм водоплавающей птице. В Японии выведена культурная форма для оформления водоемов, на соцветиях этих растений белые махровые цветки в поперечнике до 2 см.
На экскурсиях нельзя путать болотное растение частуху со стрелолистом и допускать, чтобы учащиеся брали в рот листья, черешки, корневища частухи или попытались сварить «на пробу» ее корневище, клубни. В отличие от стрелолиста частуха ядовита!
Гигрофила – род тропических прибрежных растений семейства акантовых. Оно влаголюбивое («гигро» – влага), но не водное. В природе обитает по берегам водоемов с колеблющимся уровнем воды, а в тропиках эти колебания достигают 5 – 20 м. Разные виды этого рода распространены в тропических зонах Азии и Америки.
Стебель тонкий, ветвится; листья супротивные, овальные, с закругленным концом у одних видов, длинные, ланцетовидные, с острым концом у других; размер листьев колеблется в зависимости от вида от 5 до 15 – 20 см.
Приспособление к широкому распространению, завоеванию новых ареалов проявляется у гигрофил в их удивительном вегетативном размножении. У этих растений не только черешок или обломок стебля может дать начало новому растению, но и мельчайшая часть листа.
В природе семена этого растения расселяют птицы и другие животные.
Используя сравнительно небольшую группу растений, учитель может существенно оживить уроки биологии (и не только по курсу ботаники), способствовать развитию интереса к предмету у учащихся, развить их способность творчески осмысливать биологические закономерности и понятия.
Дата добавления: 2016-01-26; просмотров: 1290;