Молекулярная природа генов с количественным эффектом
Здесь мы коснемся сложного вопроса, еще далекого от полной ясности, однако весьма немаловажного для последующего изложения, – молекулярной организации наследственного вещества. Хромосома многоклеточных – это очень длинная (до нескольких сантиметров) единая молекула ДНК. Хотя она и является носителем наследственной информации, из этого совсем не следует, что любая нуклеотидная последовательность генома обязательно влияет на приспособленность. Анализ строения ДНК многоклеточных показывает, что генетическая информация (по крайней мере, доступная нашему пониманию) размещена вдоль этой гигантской молекулы далеко не равномерно. Более того, нет сомнений, что гены занимают в ней только небольшую часть. При этом сам ген состоит из совершенно разных по информационному содержанию сегментов – экзонов и интронов. Первые чаще всего кодируют первичную структуру какого‑нибудь белка, а вторые обычно ничего не кодируют (рис. 26).
В межгенных промежутках (а иногда и в интронах) разбросаны нуклеотидные последовательности, имеющие отношение к генной регуляции. Активность гена проявляется в производстве им своего слепка – молекулы РНК, из которой затем вырезаются участки, соответствующие интронам. Заметим, что транскрипция РНК является лишь одним из этапов экспрессии гена. Другие этапы включают в себя синтез, сборку и созревание белковых молекул, а также их транспорт и встраивание в соответствующие субклеточные структуры.
В многоклеточном организме большое число генов находится под жестким контролем программ развития всевозможных морфологических структур, и многие из них активны только в клетках определенного типа. Многочисленные данные, полученные в последнее время, свидетельствуют, что такой контроль осуществляется согласованным взаимодействием регуляторных белков (транс‑факторов) с довольно короткими и весьма специфичными последовательностями ДНК (цис‑элементами). Большинство цис‑элементов расположено невдалеке от точки начала транскрипции гена; одни из них ответственны за тканеспецифичность генной экспрессии, другие – за ее уровень.
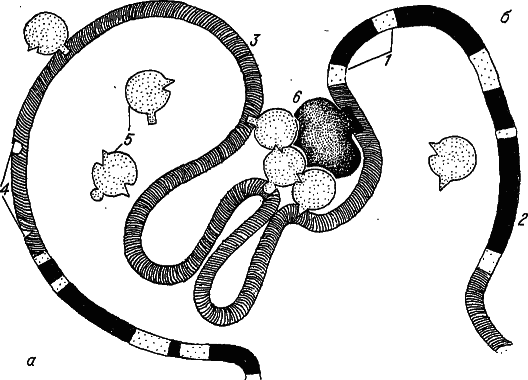
Рис. 26. Строение сегмента хромосомы эукариот. а – неактивный, б – активный ген. 1 – экзоны; 2 – интроны; 3 – межгенные участки; 4 – цис‑элементы; 5 – транс‑факторы; 6 – РНК‑полимераза.
Для включения гена (т. е. для инициации транскрипции РНК) необходимо образование комплекса транс‑факторов с белками транскрипционной машины (представленной прежде всего субъединицами РНК‑полимеразы). При этом цис‑элементы сближаются, а отрезки ДНК между ними сгибаются в петли. Таким образом, большой сегмент хромосомы с геном и протяженными (до нескольких тысяч нуклеотидов) соседними последовательностями выступает как единая матрица для сборки мультимолекулярного ансамбля, инициирующего транскрипцию. Понятно, что изменение числа цис‑элементов и даже изменение расстояния между ними может отразиться на скорости сборки такого ансамбля, т. е. в конечном счете на уровне генной экспрессии.
Хромосомы многоклеточных буквально начинены так называемыми мобильными элементами – всевозможными последовательностями ДНК, способными к независимому «размножению» и передвижению внутри генома. По‑видимому, именно эти перемещения ответственны за возникновение большей части так называемых спонтанных мутаций, составляющих основу генетической изменчивости природных популяций. Легко представить, что встраивание довольно крупного отрезка чужеродной ДНК в экзонный участок гена приведет к инактивации последнего, а соответствующая мутация пополнит ряды рецессивных леталей, подлежащих удалению очищающим отбором. Однако гораздо чаще мобильные элементы должны попадать в протяженные межгенные промежутки, что может отразиться лишь на экспрессии расположенных по соседству генов. На уровне фенотипа такие изменения наследственного материала будут выглядеть скорее всего как полигенные мутации.
Мукаи проследил за уровнем экспрессии гена алкогольдегидрогеназы (фермента, восстанавливающего спирт) у дрозофилы в течение 300 поколений. Хромосома с этим геном была выведена из‑под контроля очищающего отбора, что позволяло проследить за накоплением спонтанных мутаций, затрагивающих активность фермента. За время наблюдения межлинейная дисперсия по ферментативной активности возросла в 27 раз, хотя среднелинейное значение активности осталось прежним. Также не изменились и физико‑химические свойства алкогольдегидрогеназы, обусловленные первичной структурой белка. Мукаи предположил, что мутации затронули какие‑то районы, расположенные за пределами гена.
Мобильность
По Дарвину, эволюция основана на отборе особей с едва заметными уклонениями фенотипа. Поскольку отбор направлен на изменение величины определенного количественного признака, принципиальное значение имеет не мутабильность генов, а мутабильность признаков. Существует несколько способов измерения последней. Например, каждая особь изогенной линии самоопыляющихся растений может стать основательницей дочерней сублинии. Если все особи‑основательницы имеют тождественные генотипы, то средние значения количественных признаков в сублиниях будут совпадать. Если же особь‑основательница несет мутацию, затрагивающую развитие одного из количественных признаков, то среднее значение данного признака у потомков мутантною индивида скорее всего будет смещено. Следовательно, определяя генотипические значения признака в большой совокупности дочерних сублиний, можно оценить уровень его мутабильности. Такого рода анализ был проведен у кукурузы и у мыши, в обоих случаях получена удивительно высокая величина мутабильности – около 0,01 мутаций на признак на геном за поколение.
Таким образом, в результате мутирования в любой крупной популяции за одно поколение должна возникать новая генетическая дисперсия по любому количественному признаку. Получается, что даже изогенная линия может отвечать на отбор. Как мы помним (см. уравнение (4.19)), величина этого ответа пропорциональна наследуемости признака, т. е. доле генетической компоненты в его фенотипической дисперсии. В экспериментах по отбору на число щетинок в изогенных линиях дрозофилы скорость появления мутационной дисперсии оценена в несколько тысячных от величины фенотипической дисперсии. Длительный и интенсивный отбор должен вести к постепенному исчерпанию запасов генетической изменчивости в популяции. Поэтому ответ на отбор в конце концов будет определяться лишь скоростью возникновения мутационной дисперсии.
Процесс мутирования по разным генам независим, поэтому в первом приближении можно считать, что мутационная дисперсия по величине признака, возникающая за поколение, равна сумме мутационных дисперсий по каждому локусу. Выходит, что с ростом генетической сложности признака должна расти и мутационная дисперсия его величины. В предыдущей главе мы установили, что увеличение числа специальных генов ведет к росту размеров и мощности рабочей структуры. В связи с этим приобретает особое значение вопрос, как будут соотноситься между собой обе характеристики структуры (мощность и мутационная изменчивость) при увеличении числа специальных генов.
В качестве меры изменчивости структуры, обусловленной мутированием одного локуса, удобно взять сигму мутационного эффекта (σ0), которую можно трактовать как наиболее вероятное значение сдвига мощности структуры при мутировании одного из ее специальных генов. Так как естественный отбор «измеряет» эффект мутационных замен логарифмической линейкой, то имеет смысл оценивать величину σ0 в долях от мощности интактной структуры (напомним, что ΔlnP ≈ ΔР/Р ).
Если бы мощность органа определялась суммой вкладов его специальных генов, то величина относительного прироста мощности при добавлении к программе развития еще одного гена была бы тем меньше, чем выше генетическая сложность органа. Очевидно, что в данном (аддитивном) случае генетического контроля накопление специальных генов вело бы к созданию рабочих структур, мало чувствительных к мутациям, так как величина сигмы мутационного эффекта уменьшалась бы с увеличением числа их специальных генов.
Однако обычно мощность органа представима в виде произведения сомножителей, олицетворяющих собой вклады отдельных специальных генов, поэтому добавление еще одного такого гена увеличивает мощность органа в некоторое число раз (вне зависимости от его мощности или от числа специальных генов), т. е. величина относительного прироста мощности рабочей структуры не зависит от ее генетической сложности. В данном (мультипликативном) случае генетического контроля увеличение числа специальных генов не снижает чувствительности органа к мутациям, поскольку любой сдвиг (в процентах) вклада отдельного гена представляет собой сдвиг на тот же процент мощности органа. Таким образом, сигма мутационного эффекта на мощность рабочей структуры не должна зависеть от числа ее специальных генов.
Начиная с Дарвина неоднократно используется метафора, что естественный отбор как бы лепит новые формы, поэтому сам собою возник вопрос о пластичности материала, с которым имеет дело эволюция. Часто высказывалось, по‑видимому, совершенно правильное мнение, что биологический материал не пластилин, что формирование структур подчиняется своим законам и т. д. Не входя в дискуссию о «законах» образования биологических структур, заметим, что с функциональной точки зрения имеет значение только их мощность. В связи с этим вместо несколько неопределенной пластичности биологического материала лучше говорить об эволюционной изменчивости мощности рабочих структур. Эту изменчивость назовем мобильностью и будем ее оценивать по скорости, с которой под действием длительного движущего отбора изменяется мощность структуры. А учитывая «логарифмические очки» естественного отбора (т. е. его «склонность» оценивать не абсолютный, а относительный сдвиг величины признака), назовем мобильностью скорость, с которой изменяется логарифм мощности рабочей структуры под действием длительного движущего отбора определенного давления. Еще раз напомним, что небольшой относительный сдвиг величины признака (измеренный в долях от его исходного значения) можно трактовать как сдвиг логарифма величины признака.
Как ответ на длительный отбор мобильность (М) должна прямо зависеть от скорости возникновения мутационной дисперсии логарифма мощности структуры, что, в свою очередь, определяется числом специальных генов (N ), их средней мутабильностью и средней дисперсией мутационного эффекта <σ02>. Поскольку мощность структуры определяется мультипликативным действием ее специальных генов, то сигма мутационного эффекта σ0, измеренная в долях от мощности интактной структуры, не зависит от числа этих генов. Кроме того, число специальных генов любой морфологической структуры довольно велико, поэтому значение <σ02> для разных структур, по‑видимому, различается незначительно. Наконец, по средней мутабильности специальных генов рабочие структуры также не могут существенно различаться. Таким образом, с достаточной точностью должно выполняться соотношение
М ~ N, (6.1)
т. е. мобильность структуры пропорциональна числу ее специальных генов.
Перевернув назад несколько страниц, можно убедиться в том, что совсем недавно мы установили почти тривиальную связь между мощностью структуры у среднестатистического вида филетической группы, с одной стороны, и числом его специальных генов – с другой (см. (5.10)). Сопоставив это соотношение с (6.1), получаем, что
<lnP > ~ <М> , (6.2)
т. е. в пределах филетической группы средневидовой логарифм мощности рабочей структуры пропорционален ее средневидовой мобильности.
Дата добавления: 2016-01-26; просмотров: 1414;