Несостоятельность ортоселекции
Выше в поиске рационального объяснения долговременных эволюционных тенденций мы остановились на двух гипотезах – ортоселекции и росте адаптируемости. Хотя ортоселекция на первый взгляд выглядит предпочтительнее (поскольку обладание более сложной структурой уже кажется благом), эта гипотеза не в состоянии объяснить феномен застоя. Если преимущества сложной лопастной линии ощущались в первые 200 млн лет эволюции аммоноидей, то они должны были сохранять свое значение и далее, но за последние 130 млн лет доля аммоноидей с довольно простой сутурой не убывала. Также трудно объяснить отсутствие прогресса в строении лопастной линии гониатитов середины карбона и цератитов конца триаса (см. рис. 22, б).
Другой сильный аргумент против ортоселекции был представлен американским палеонтологом Дж. Цисне, проследившим за ростом дифференциации конечностей у водных членистоногих. Для количественной оценки этой формы морфофункционального прогресса Цисне предложил использовать степень тагмозиса конечностей (h), которую можно вычислить по формуле (2.1). Каждый отряд водных членистоногих (от примитивных трилобитов до высших ракообразных) был охарактеризован величиной h, после чего было проанализировано, как в течение 600 млн лет эволюции этих беспозвоночных изменялся средний для отряда уровень тагмозиса.
Оказалось, что в ходе исторического развития водных членистоногих величина <h> возрастала, следуя S‑образной (логистической) кривой (рис. 24). В течение первых 250 млн лет шло довольно быстрое увеличение среднего тагмозиса, затем этот процесс стал тормозиться и практически прекратился к началу мезозоя. Так что за последние 150 млн лет уровень <h> не повышался, хотя одни отряды вымирали, а другие возникали.
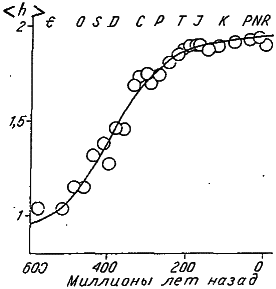
Рис. 24. Рост среднего тагмозиса (<h>) конечностей водных членистоногих в историческом развитии (по: [Cisne, 1974]).
E – кембрий, О – ордовик, S – силур, Р – палеоген, N – неоген, R – современность. Остальные условные обозначения см. рис. 22.
Данное явление нельзя объяснить исчерпанием возможностей к прогрессивной дифференциации конечностей, так как средний тагмозис на стадии плато (<h> = 1,8) намного ниже, чем у десятиногих ракообразных (h = 2,5). Низкий уровень плато объясняется существованием в течение многих миллионов лет практически постоянной доли «примитивных» отрядов. Это плато продолжительностью в 150 млн лет явно свидетельствует против таинственных внутренних (гомогенетических) факторов, толкавших членистоногих по пути прогресса, но, самое главное, оно ставит серьезные трудности и перед сторонниками ортоселекции. Ведь выгода от функциональной дифференциации конечностей, казалось бы, очевидна. Тогда почему же доля примитивных отрядов не убывала в течение последних 150 млн лет?
В 1949 г. Т. Уэстолл предпринял попытку измерить темп прогрессивной эволюции у двоякодышащих рыб. В данном случае уровень организации оценивался числом «прогрессивных» структурных новшеств (числом баллов). Динамика прогресса структурной сложности имеет тот же S‑ образный вид: период быстрого увеличения числа баллов продолжительностью около 150 млн лет сменяется гораздо более долгим плато (рис. 25).

Рис. 25. Увеличение числа прогрессивных признаков (в баллах) в историческом развитии двоякодышащих рыб (по: [Рефф, Кофмен, 1986]).
Характер кривых, передающих три случая морфологического прогресса (рост тагмозиса у членистоногих, сложности лопастной линии у аммоноидей и продвинутости организации у двоякодышащих рыб) весьма сходен. Во всех случаях первоначальный период подъема сменяется периодом застоя. Объяснение этому удивительному феномену мы попытаемся дать в следующей главе.
Дата добавления: 2016-01-26; просмотров: 857;