ПРОПРИОЦЕПТИВНЫЙ АНАЛИЗАТОР
Проприоцептивный анализатор специализируется на восприятии информации о состоянии опорно-двигательного аппарата и косвенно участвует в координации движений. Рецепторная часть этого анализатора заложена в опорно-двигательном аппарате и представлена дендритами чувствительных нейронов, оплетающими интрафузальные мышечные волокна в скелетных мышцах, либо многократно разветвляющимися между коллагеновыми или эластическими волокнами связок и сухожилий, либо входящими в состав телец Фатера-Пачини в капсулах суставов. Проприорецепторыобеспечивают восприятиеизменения длины и скорости сокращения скелетных мышц (мышечные веретена), степени растяжения сухожилий (тельца Гольджи), а также изменения давления в капсулах суставов (тельца Фатера-Пачини).Информация от рецепторов опорно-двигательного аппарата туловища и конечностей проходит через спинальные ганглии в спинной мозг, а от опорно-двигательного аппарата головы и шеи – через гассеров узел тройничного нерва, коленчатый узел лицевого нерва и каменистый узел языкоглоточного нерва в головной мозг (к чувствительным ядрам этих нервов, заложенным в ромбовидной ямке). Аксоны нейронов спинальных ганглиев, проводящие проприоцептивную информацию в направлении коры больших полушарий, следуют через спинной мозг транзитом в составе путей Голя и Бурдаха и достигают продолговатого мозга, где переключаются на нейроны ядер Голя и Бурдаха. Аксоны же нейронов этих ядер вместе с волокнами чувствительных ядер черепномозговых нервов, имеющих отношение к проведению проприоцептивной информации от головы и шеи, восходят к таламусу (латеральная специфическая группа ядер и вентральная неспецифическая группа ядер) в составе медиальной петли. Нейроны таламических бугров посылают свои проекции к корковому представительству проприоцептивного анализатора, заложенному в теменной доле больших полушарий (постцентральная извилина и верхняя теменная долька), часть волокон таламических нейронов направляется непосредственно к моторной коре (пердцентральная извилина и околоцентральная долька).
Периферическая часть проприоцептивного анализатора заложена вопрно-двигательном аппарате(секлетные мышцы, связки, сухожилия, капсулы суставов).Несмотря на существующее разнообразие рецепторов опрно-двигательного аппарата все они с функциональной точки зрения являются первичночувствующими механорецепторами.
Рецепторы скелетных мышц – мышечные веретена (реагируют собственно на изменение длины мышцы, а также на скорость изменения ее длины)
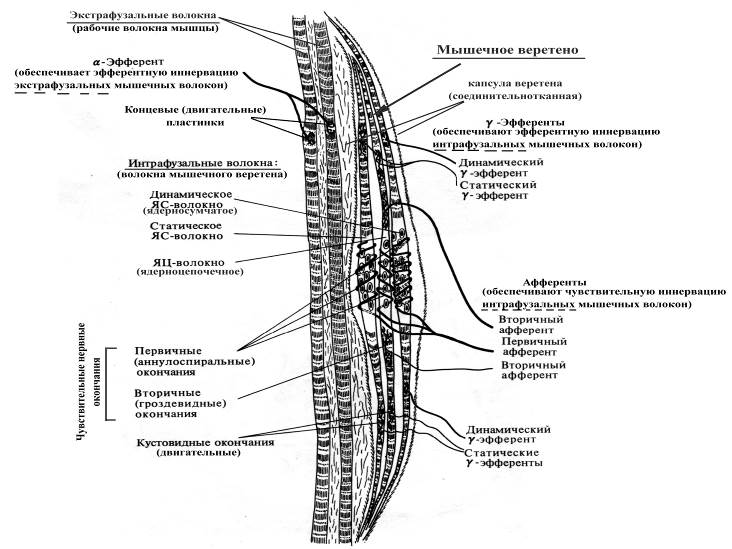
Рис. 16. Схема строения мышечного веретена
Длина мышечных веретен составляет около 3-5 мм, толщина ~0,2 мм, ориентированы в мышцах продольно (в связи с этим при растяжении мышцы растягивается и веретено). Каждое веретено содержит от 2 до 12 интрафузальных мышечных волокон двух типов:
Ø ядерно-сумчатые (в состав веретена входит от 1 до 4 таких волокон, толще и длиннее ядерно-цепочечных, ядра образуют скопления в центре волокна), получают афферентную иннервацию только из одного источника (т.н. первичный афферент, спирально закручиваясь, оплетает ядерную сумку);
Ø ядерно-цепочечные (в составе веретена насчитывается до 10 таких волокон, тоньше и короче ядерно-сумчатых, ядра расположены под сарколеммой в виде цепочки на всем протяжении волокна), получают афферентную иннервацию из двух источников – первичный афферент (более толстый, чем вторичный) оплетает ядерно-цепочечное волокно по центру, а вторичный афферент оплетает ядено-цепочечное волокно по одному с каждой стороны от нервных окончаний первичного афферента.
Интрафузальные мышечные волокна ориентированы параллельно экстрафузальным, в связи с чем при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. Адекватным раздражителем для чувствительных нервных волокон, оплетающих интрафузальные мышечные волокна, является растяжение мышцы, тогда как ее укорочение деактивирует афференты мышечных веретен.
В расслабленной мышце афферентная импульсация от веретен очень слабая, тогда как при растяжении мышцы она резко возрастает. Причем первичные афференты, оплетающие интрафузальные мышечные волокна, реагируют на степень и на скорость растяжения мышцы (динамический ответ), а вторичные, оплетающие ядерно-цепочечные интрафузальные волокна, – только на степень растяжения мышцы (статический ответ).
Наряду с афферентной иннервацией оба типа волокон мышечных веретен получают эфферентную иннервацию со стороны g-мотонейронов. Физиологическая роль эфферентной иннервации интрафузальных мышечных волокон состоит в инициации сокращения их периферических частей, и как следствие, усилении натяжения, а значит и повышении чувствительности на растяжение при любой данной длине мышцы. Кроме того, постоянная базальная активность g-мотонейронов способствует поддержанию в постоянном тонусе интрафузальных мышечных волокон, что обеспечивает постоянное возбуждение афферентов мышечных веретен и как следствие, a-мотонейронов, получающих информацию от этих афферентов и иннервирующих экстрафузальные (рабочие) волокна мышцы. Тоническая же активность a-мотонейронов сопровождается поддержанием постоянного (базального) тонуса всей скелетной мышцы. Наряду с описанной зависимостью между возбуждением g- и a-мотонейронов, имеет место и обратная a-g-коактивация. Причем уровень возбуждения g-системы тем выше, чем интенсивнее возбуждены a-мотонейроны данной мышцы, т. е, чем больше сила сокращения.
 Само по себе сокращение мышцы сопровождается дезактивацией афферентов мышечных веретен (они активируются в ответ на растяжение мышцы), но возбуждение a-мотонейронов приводит к сочетанной активации и g-мотонейронов, вызывающих сокращение интрафузальных мышечных волокон и, как следствие, активацию первичных и вторичных афферентов. Следовательно, состояние афферентов мышечных веретен в активной мышце является результатом двух противоположных факторов: степени укорочения мышцы (дезактивирующей афференты веретен) и состояния g-системы, активирующейся сочетанно с возбуждением a-мотонейронов и вызывающей повышение импульсации в афферентах мышечных веретен. Во время активных движений актвированные a-мотонейронами g-мотонейроны, инициируя сокращение интрафузальных мышечных волокон, поддерживают постоянную активность афферентов мышечных веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на вызванные помехами неравномерности движения как увеличением, так и уменьшением частоты им пульсации и участвовать таким образом в коррекции движений.
Само по себе сокращение мышцы сопровождается дезактивацией афферентов мышечных веретен (они активируются в ответ на растяжение мышцы), но возбуждение a-мотонейронов приводит к сочетанной активации и g-мотонейронов, вызывающих сокращение интрафузальных мышечных волокон и, как следствие, активацию первичных и вторичных афферентов. Следовательно, состояние афферентов мышечных веретен в активной мышце является результатом двух противоположных факторов: степени укорочения мышцы (дезактивирующей афференты веретен) и состояния g-системы, активирующейся сочетанно с возбуждением a-мотонейронов и вызывающей повышение импульсации в афферентах мышечных веретен. Во время активных движений актвированные a-мотонейронами g-мотонейроны, инициируя сокращение интрафузальных мышечных волокон, поддерживают постоянную активность афферентов мышечных веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на вызванные помехами неравномерности движения как увеличением, так и уменьшением частоты им пульсации и участвовать таким образом в коррекции движений.
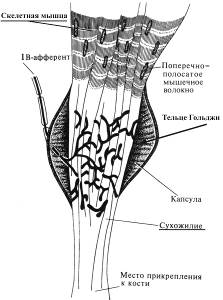
Рецепторы связок и сухожилий – тельца Гольджи (реагируют на растяжение связок и сухожилий)
Имеют строение, подобное тельцам Руффини соединительной ткани кожи, но отличаются большими размерами. Находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам.
Сухожильные рецепторы Гольджи слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении; интенсивность их импульсации пропорциональна силе сокращения мышцы, что позволяет рассматривать их как источник информации о силе, развиваемой мышцей. На спинальном уровне афферентные нервные волокна, идущие от рецепторов Гольджи, вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антогониста.
Рецепторы капсул сустав – тельца Фатера-Пачини (рецепторы, реагирующие на изменение давления в капсуле сустава, а следовательно, на его положение или изменение суставного угла). Подобно проприорецепторам скелетных мышц и тельцам Гольджи участвуют в системе обратных связей от двигательного аппарата.
ОБОНЯТЕЛЬНЫЙ АНАЛИЗАТОР
Обонятельный анализатор человека специализируется на восприятии запахов молекул пахучих веществ, попадающих с вдыхаемым воздухом в полость носа и носоглотку. Человек по способности различать запахи относится к микросоматикам (обонятельный анализатор у человека не играет столь важного значения для ориентации в окружающем мире, как, например, у многих ползающих животных (крысы, мыши и другие мелкие грызуны) или у большинства насекомоядных, жвачных, копытных и хищных животных), относящихся к макросоматикам. Его периферическая часть заложена в обонятельном эпителии слизистой носовой полости и представлена собственно дендритами обонятельных нейронов (первичночувствующие хеморецепторы), аксоны которых образуют обонятельный нерв (Iп), направляющийся непосредственно к большому (конечному) мозгу. Обонятельная информация является единственной чувствительной информацией, следующей напрямую к большому мозгу, минуя таламические бугры (коллекторы всей чувствительной информации). Волокна обонятельного нерва, не перекрещиваясь, переключаются на нейроны обонятельной луковицы (прилежит к медиальной поверхности лобных долей больших полушарий). Аксоны же нейронов обонятельной луковицы формируют обонятельный тракт, волокна которого могут следовать к переднему обонятельному ядру, обонятельному бугорку, ядрам миндалевидного комплекса (структуры т.н. обонятельного мозга). Аксоны нейронов этих подкорковых обонятельных центров направляются в парагиппокампальную извилину и крючок (корковый обонятельный центр). Причем большинство областей проекции обонятельного тракта не являются необходимыми для опознания запахов, а служат скорее ассоциативными зонами, обеспечивающими связь обонятельной системы с другими сенсорными системами и организацию на основе этой связи ряда сложных форм поведения (пищевой, оборонительной, половой и т.д.).
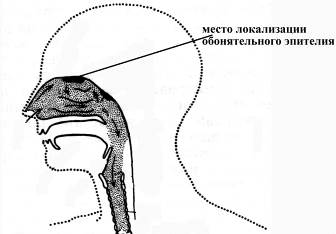
Обонятельный эпителий выстилает большую часть крыши каждой полости носа, начинается спереди от переднего конца верхней носовой раковины и простирается назад приблизительно на 1 см; с крыши каждой полости носа он спускается по обеим ее сторонам (причем по медиальной поверхности (носовой перегородке) – приблизительно на 1 см книзу, а по латеральной (крылу носа) вниз почти по всей поверхности носовой раковины).
Рис. 17. Локализация обонятельного эпителия в слизистой носовой полости человека
В обонятельном эпителии различают следующие типы клеток:
Ø собственно обонятельными нейронами, дендриты которых достигают апикальной поверхности эпителиального пласта, на уровне которой заканчиваются обонятельной булавой, от которой отходят 6-12 ресничек (длиной до 10мкм). Считают что в роли обонятельного рецептора, возбуждающегося в результате контакта с молекулами пахучих веществ, выступает обонятельная булава, а отходящие от нее реснички, с одной стороны, увеличивают площадь поверхности булавы, а с другой, могут участвовать в активном захвате молекул пахучих веществ.
Ø малодифференцированными (стволовыми) нейронами, являющимися источником обновления популяции обонятельных нейронов,
Ø поддерживающими (опорными) эпителиоцитами,представляющими собойклетки цилиндрической формы, на апикальном конце которых имеется большое количество микроворсинок увеличивающих площадь их контакта с молекулами пахучих веществ и соответственно концентрацию молекул пахучих веществ в области дендритных разветвлений обонятельных нейронов.
Толщина обонятельного эпителия составляет 100-150мкм, а средний диаметр тел обонятельных нейронов – 5-10мкм. Общее количество обонятельных нейронов у человека достигает около 10млн. В слизистой носа, покрытой обонятельным эпителием, имеются боуменовы железы, секрет которых увлажняет поверхность обонятельных нейронов и поддерживающих эпителиоцитов, способствуя более длительному удержанию молекул пахучих веществ в области обонятельных рецепторов.
Активация обонятельных рецепторов в результате контакта с молекулами пахучих веществ согласно современным представлениям происходит следующим образом. Молекулы пахучих веществ, вступая в контакт с обонятельной булавой, взаимодействуют со специализированными белками, встроенными в мембрану рецептора, в результате чего изменяется проницаемость мембраны для определенных веществ, что приводит к генерации рецепторного потенциала, вызывающего импульсное возбуждение мембраны аксонного холмика обонятельного нейрона. Причем каждый обонятельный рецептор отвесчает не на один, а на многие пахучие вещества, отдавая «предпочтение» некоторым из них. считают, что на этих свойствах рецепторов, различающихся по своей настройке на разные группы веществ, может быть основано кодирование информации о запахах (пространственное кодирование) и их опознание в центрах обонятельного анализатора.
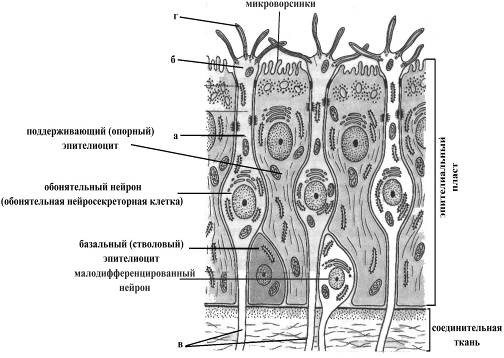
Рис. 18. Схема ультраструктур-ной организации обонятельного эпителия
а-дендрит обонятельного ней-рона
б-булавовидная терминаль ден-дрита
в-аксоны обонятельных нейрон-нов, формирующие обонятель-ный нерв, Iп.
г-реснички
Чувствительность обонятельного анализатора человека чрезвычайно высока: один обонятельный рецептор может быть возбужден одной молекулой пахучего вещества, а возбуждение небольшого количества рецепторов приводит к формированию ощущения определенного запаха. Вместе с тем способность человека различать запахи по своей интенсивности довольна низкая: наименьшее воспринимаемое различие в силе запаха составляет 30-60% от его исходной концентрации.
Адаптация в обонятельном анализаторе происходит сравнительно медленно (десятки секунд или минуты) и зависит от скорости потока воздуха над обонятельным эпителием и концентрации пахучего вещества.
ВКУСОВОЙ АНАЛИЗАТОР
Вкусовой анализатор специализируется на восприятии и обработке вкусовых раздражений, действующих на вкусовые рецепторы, заложенные в слизистой проксимальных отделов пищеварительной трубки. Вкусовые рецепторы воспринимают информацию о характере и концентрации веществ, поступающих в рот. Их возбуждение запускает сложную цепь реакций разных отделов мозга, приводящих к различной работе органов пищеварения или удалению вредных для организма веществ, попавших в ротовую полость.
Периферическая часть вкусового анализатора локализована в эпителиальном пласте слизистой начальных отделов пищеварительной трубки и представлена вкусовыми рецепторными клетками (входят в состав вкусовых луковиц (или почек)), вступающими в синаптический контакт с дендритами чувствительных нейронов. Тела чувствительных нейронов, имеющих отношение к проведению чувствительной информации от вкусовых рецепторных клеток в центральную нервную систему, заложены в чувствительных узлах лицевого (коленчатый узел), языкоглоточного (каменистый узел) и блуждающего (яремный узел) нервов. Причем информация от вкусовых рецепторов передних 2/3 языка проводится в центральную нервную систему через коленчатый узел лицевого нерва, от задней 1/3 языка – через каменистый узел языкоглоточного нерва, а от задней части глотки и надгортанника – через яремный узел блуждающего нерва. Аксоны чувствительных вкусовых нейронов, имеющих отношение к проведению вкусовой информации, входят в головной мозг в составе чувствительных ветвей лицевого, языкоглоточного и блуждающего нервов и переключаются на нейроны солитарного ядра (ядра одиночного пути), заложенного в ромбовидной ямке, и являющегося общим чувствительным ядром для этих трех пар черепномозговых нервов. Аксоны же нейронов солитарного ядра восходят к таламическим буграм в составе медиальной петли и переключаются на нейроны латеральной группы ядер, дающих проекции к вкусовой коре (корковое представительство вкусового анализатора находится в височной доле, вблизи латеральной борозды).
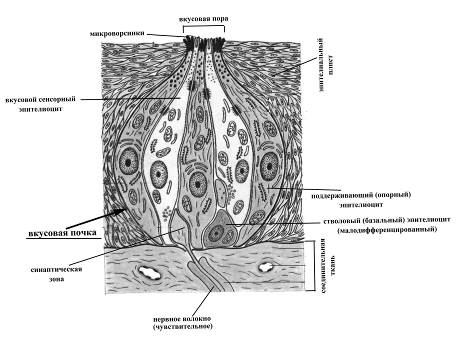 Периферическая часть вкусового анализатора представлена вкусовыми рецепторными клетками, входящими в состав вкусовых почек или луковиц. Вкусовые почкизаложены в эпителии боковых поверхностей сосочков языка (кроме нитевидных), слизистой мягкого неба, миндалин, задней части глотки и верхней поверхности надгортанника. Причем апикальная поверхность луковиц достигает поверхности эпителиального пласта. Общее их количество у человека составляет около 10000.
Периферическая часть вкусового анализатора представлена вкусовыми рецепторными клетками, входящими в состав вкусовых почек или луковиц. Вкусовые почкизаложены в эпителии боковых поверхностей сосочков языка (кроме нитевидных), слизистой мягкого неба, миндалин, задней части глотки и верхней поверхности надгортанника. Причем апикальная поверхность луковиц достигает поверхности эпителиального пласта. Общее их количество у человека составляет около 10000.

Рис. 19. Листовидные сосочки языка Рис. 20. Схема ультрастуктурной организации вкусовой почки
Вкусовые луковицы имеют колбовидную форму, на их вершине, обращенной к поверхности эпителиального пласта, находится вкусовая пора – отверстие, через посредство которого молекулы различных веществ поступают внутрь почки. Диаметр вкусовых луковиц составляет в среднем 70мкм, они включают два типа клеток: вкусовые (рецепторные) и поддерживающие (опорные). В каждой луковице содержится 2-6 рецепторных клеток, на апикальной поверхности которых имеются микроворсинки (диаметр 0,1-0,2мкм, длина 1-2мкм), увеличивающие площадь поверхности клетки, а следовательно, и площадь ее контакта с молекулами веществ, поступающих в луковицу. Более того, предполагают, что именно в области микроворсинок расположены активные центры – стериоспецифические участки рецептора, избирательно воспринимающие адсорбцию разных веществ и играющие определяющую роль в процессе возбуждения вкусовой рецепторной клетки. Вкусовые клетки характеризуются очень высокой скоростью физиологической регенерации: средняя продолжительность жизни вкусовых эпителиоцитов составляет 250ч (около 10суток), после чего они сменяются молодыми, движущимися к центру вкусовой почки от ее периферии. Базальная поверхность рецепторных вкусовых клеток вступает в синаптичесий контакт с дендритами чувствительных нейронов, тела которых могут быть заложены в чувствительных узлах лицевого (коленчатый узел), языкоглоточного (каменистый узел) или блуждающего (яремный узел) нервов. Аксоны чувствительных нейронов в составе чувствительных ветвей указанных нервов направляются в центральную нервную систему. Следовательно, вкусовые рецепторы являются вторичночувствующими хеморецепторами. С функциональной точки зрения различают 4 типа вкусовых рецепторов, проявляющих наибольшую чувствительность к определенным вкусовым качествам: горькому, соленому, сладкому или кислому. Различные типы вкусовых рецепторов неравномерно распределены на поверхности языка. Так, наибольшая плотность рецепторов, проявляющих максимальную чувствительность к кислому, характерна для латеральных поверхностей языка, сладкому – его кончика, горькому – корня, а рецепторы, наиболее чувствительные к соленому, встречаются по всей поверхности языка. Предполагают, что информация о четырех основных вкусовых ощущениях кодируется не частотой импульсации в одиночных волокнах, а разным распределением частоты разрядов в большой группе волокон, одновременно, но по-разному, возбуждаемых вкусовым веществом.
Вкусовой анализатор подобно другим обладает способностью к адаптации. Причем продолжительность адаптации пропорциональна концентрации раствора вещества, имеющего определенное вкусовое качество, и отчасти зависит от характера вкусового воздействия. Так, адаптация к сладкому и соленому развивается быстрее, чем к горькому и кислому. Возможна и перекрестная адаптация (изменение чувствительности к одному веществу при действии другого). В частности, адаптация к сладкому повышает чувствительность к веществам, имеющим все остальные вкусовые качества.
В естественных условиях вкусовые ощущения комбинируются с обонятельными, тактильными и термическими, также создаваемыми пищей, поступающей в ротовую полость.
Дата добавления: 2016-01-07; просмотров: 1863;