ВЕСТИБУЛЯРНЫЙ АНАЛИЗАТОР
Вестибулярный анализатор специализируется на восприятии изменений положения тела (прежде всего головы) в пространстве, а также действия на организм ускорений при движении и играет важную роль, наряду со зрительным и проприоцептивным анализаторами, в пространственной ориентировке человека. В условиях покоя или равномерного движения рецепторы вестибулярного анализатора не возбуждаются.
Периферическая (рецепторная) часть вестибулярного анализатора находится во внутреннем ухе (перепончатый лабиринт преддверия и полукружных каналов) и представлена вестибулярными волосковыми клетками, вступающими в синаптический контакт с дендритами вестибулярных нейронов, тела которых находятся в вестибулярном ганглии (заложен в толще пирамидки височной кости). Аксоны вестибулярных нейронов образуют преддверную (вестибулярную) ветвь преддверно-улиткового нерва (VIIIп) и направляются к вестибулярным ядрам, локализованным в латеральных углах ромбовидной ямки (латеральное вестибулярное ядро (ядро Дейтерса), медиальное вестибулярное ядро (ядро Швальбе), верхнее вестибулярное ядро (ядро Бехтерева) и нижнее вестибулярное ядро (ядро Ролкера)). Нейроны вестибулярных ядер, в свою очередь, дают проекции к
Ø спинному мозгу (от латерального и нижнего вестибулярных ядер начинается вестибуло-спинальный путь),
Ø ретикулярной формации ствола головного мозга,
Ø мозжечку,
Ø красному ядру и черной субстанции среднего мозга,
Ø вегетативным ядрам,
Ø а также к нейронам латеральной группы ядер таламических бугров, посылающих свои аксоны к коре больших полушарий (задняя часть постцентральной извилины и нижняя часть моторной зоны, локализованной в предцентральной извилине и центральной борозде).
Связь нейронов вестибулярных ядер со спинным мозгом и со структурами ствола головного мозга обеспечивает возможность быстрого (неосознанного) перераспределения тонуса определенных групп скелетных мышц, а также изменения деятельности некоторых внутренних органов в ответ на возбуждение ветибулорецепторов. Высший же анализ информации, поступающей от вестибулорецепторов, происходит на уровне коры больших полушарий и заканчивается формированием у человека представления о положении своего тела и головы в пространстве.
Вестибулорецепторы заложены в перепончатом лабиринте преддверия (маточке и мешочке) и ампул (начальных расширений) полукружных каналов внутреннего уха, формирующих в своей совокупности вестибулярный аппарат. В маточке и мешочке сенсорные образования имеют форму пятна (макулы), над которым нависает желатинозная отолитовая мембрана, включающая кристаллы бикарбоната кальция (удельный ее вес в 3 раза больше такового эндолимфы, заполняющей полость перепончатого лабиринта внутреннего уха). В ампулообразных расширениях перепончатых полукружных каналов рецепторные структуры вестибулярного анализатора имеют вид поперечного гребня (кристы), над поверхностью которого расположена желатинозная мембрана – купула, лишенная включений бикарбоната кальция (в связи с чем ее удельный вес такой же, как и эндолимфы, заполняющей перепончатый лабиринт полукружных каналов). Причем если отолитовая мембрана лежит над пятнами маточки и мешочка наподобие плоского камня над пластиной, то купула, подобно створчатому клапану или шарнирной дверце, идет от гребешка прямо поперек ампулы, замыкая ее просвет.
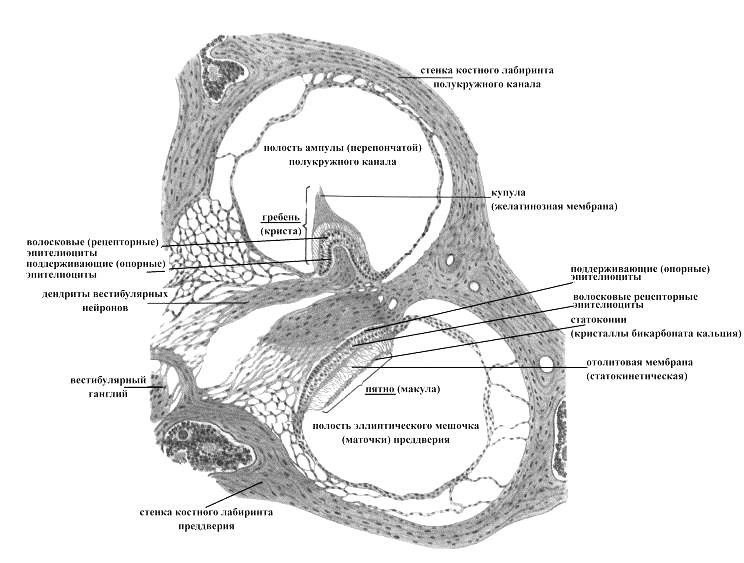
Рис. 13. Вестибулярный аппарат (поперечный разрез через пятно эллиптического мешочка и ампулу полукружного канала)
Макулы (пятна) маточки и мешочка преддверия и гребни (кристы) ампул перепончатых полукружных каналов состоят из утолщенного эпителия, включающего волосковые (рецепторные) и поддерживающие (опорные) клетки. Волосковые клетки по форме могут быть цилиндрическими и колбовидными. На апикальной поверхности этих клеток расположены стереоцилии, склеенные между собойи неподвижные (60-80 штук, производные мембраны, называемые также волосками или микроворсинками) и, как правило, одна ресничка – киноцилия (длинная и подвижная). Базальная поверхность волосковых клеток принимает участие в образовании синаптического контакта с дендритами вестибулярных нейронов.
Волосковые клетки макул перепончатого лабиринта преддверия воспринимают действие силы тяжести и линейных ускорений на организм. Если голова, а следовательно, и макула наклонены относительно силы тяжести, то отолитовая мембрана также стремится скользнуть наклонно, при этом тянет волоски волосковых клеток, деформация которых приводит к возбуждению волосковых клеток. Волосковые клетки гребней ампул перепончатых полукружных каналов выявляют изменения углового ускорения. Когда голове придано угловое ускорение, эндолимфа в определенном полукружном канале стремится сохранить неподвижность в силу инерции, а перепончатые стенки движутся относительно нее. Это движение смещает купулу из ее обычного положения относительно гребешка, смещенная же купула тянет волоски волосковых клеток, деформация которых приводит к возбуждению волосковых клеток (генерации рецепторного потенциала). Возбуждение волосковых клеток, в свою очередь, сопровождается выделением в синаптическую щель из их пресинаптических окончаний медиатора. Медиатор же, достигая постисинаптической мембраны (участка мембраны дендрита вестибулярного нейрона), вызывает ее деполяризацию (возникновение постсинаптического потенциала), порождающую нервный импульс в соседних участках мембраны вестибулярного нейрона. Следовательно, вестибулярные рецепторы представляют собой в функциональном плане вторичночувствующие механорецепторы.
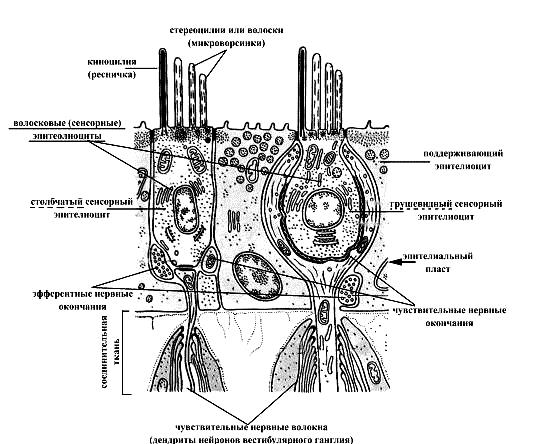
Рис. 14. Схема ультрамикроскопического строения вестибулярных рецепторов
Чувствительность вестибулярного анализатора здорового человека очень высока. Так, отолитовый аппарат позволяет воспринимать наклон головы в сторону всего на 1º, а рецепторная система полу кружных каналов – замечать ускорения вращения всего в 2-3 º/с2.
КОЖНЫЙ АНАЛИЗАТОР
Кожный анализатор специализируется на восприятии воздействий, оказываемых на кожу человека, и характеризуется очень широким рецептивным полем (от 1,4 до 2,1м2). Чувствительная информация от кожи туловища и конечностей проводится нейронами спинальных ганглиев в спинной мозг, а от кожи головы и шеи – нейронами полулунного узла тройничного нерва (Vп) в головной мозг (к чувствительным ядрам этого нерва, заложенным в ромбовидной ямке). Аксоны нейронов спинальных ганглиев, доставляющие информацию от болевых и температурных рецепторов кожи туловища и конечностей, переключаются на нейроны собственного ядра задних рогов спинного мозга, тогда как информация от тактильных рецепторов кожи туловища и конечностей проводится нейронами спинальных ганглиев к желатинозной субстанции Ролланда. Аксоны нейронов собственного ядра задних рогов спинного мозга и желатинозной субстанции Ролланда восходят к головному мозгу, формируя соответственно латеральный и вентральный спинно-таламические тракты, которые перекрещиваются еще на уровне спинного мозга. Нервные волокна этих двух трактов входят в головной мозг и вместе с аксонами нейронов чувствительных ядер тройничного нерва, проводящих информацию от рецепторов кожи головы и шеи, следуют в мозговом стволе в составе медиальной петли и достигают таламических бугров (латеральной специфической группы ядер и вентральной неспецифической группы ядер). Аксоны же нейронов специфических ядер таламуса, имеющих отношение к обработке кожной информации, восходят к корковому представительству кожного анализатора, заложенному в теменной доле коры (постцентральная извилина и верхняя теменная долька).
Рецепторная часть кожной сенсорной системы заложена в толще кожи: в эпителиальном пласте, сосочковом и сетчатом слоях дермы, подкожной жировой клетчатке. В функциональном плане чувствительные нервные окончания кожи можно классифицировать на три группы:
Ø тактильные (воспринимают прикосновение, давление, вибрацию)
Ø температурные
ü холодовые (в среднем насчитывается в коже 250тыс., заложены более поверхностно по сравнению с тепловыми (на глубине 0,17мм), максимально возбуждаются при температурах от 10 до 41ºС)
ü тепловые (в среднем насчитывается в коже 30 тыс., заложены более глубоко по сравнению с холодовыми (на глубине 0,3мм), максимально возбуждаются при температурах от 20 до 50ºС)
Ø болевые (ноцицепторы).
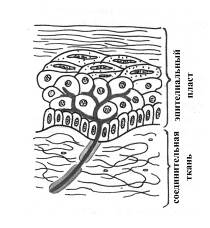 В морфологическом плане рецепторы кожи отличаются большим разнообразием, локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. Кожные рецепторы могут быть представлены следующими морфологическими типами:
В морфологическом плане рецепторы кожи отличаются большим разнообразием, локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. Кожные рецепторы могут быть представлены следующими морфологическими типами:
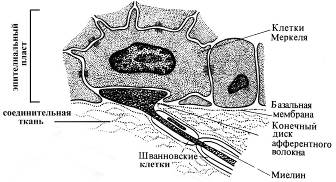
Ø Свободные нервные окончания (разветвления дендритов чувствительных нейронов)
Типичны для эпителиального пласта кожи и роговицы глаза, где могут выступать в качестве механорецепторов (тактильные рецепторы и ноцицепторы) и терморецепторов.
Ø Свободные нервные окончания, оплетающие волосяные луковицы
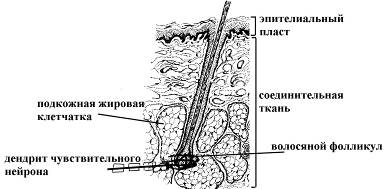 |
Типичны для соединительной ткани кожи, имеющей волосяной покров, выступают в качестве механорецепторов (тактильных рецепторов, реагирующих на прикосновение и давление).
Дата добавления: 2016-01-07; просмотров: 1845;