Проблема сохранения биотического разнообразия
Естественно, что чрезмерное потребление и уничтожение биоты человеком обуславливает в качестве одной из главнейших проблем глобальной экологии вопросы сохранения биотического разнообразия.
Выделяют три иерархических уровня биоразнообразия: генный, видовой и экосистемный. Генный уровень чрезвычайно велик и пока еще слабо исследован. Видовое разнообразие все еще оценивается весьма неоднозначно. Общее количество видов находится в пределах от 3,6 до 112 млн, в том числе насекомых от 2 до 100 млн (по данным различных исследователей). Согласно последней авторитетной оценке ЮНЕП (1995), наиболее вероятное количество видов - 13-14 млн, но описано к настоящему времени всего 1,75 млн, что едва составляет 13%.
Экосистемный уровень образует в рангах биоразнообразия следующий ряд: влажные тропические леса, коралловые рифы, сухие тропические леса, влажные леса умеренного климата, океанические острова, обстановки средиземноморского климата, безлесные ландшафты.
Фиторазнообразие в общем определяет биопродуктивность и биоразнообразие. Так, наибольшая первичная продуктивность как раз и характерна для влажных лесов экваториального пояса (примерно 4000 т/км2/год). Субтропические леса производят 2000 т/км2, а тайга 700 т/км2.
Высшие растения мира насчитывают вероятно 400000 видов и от них зависят порядка 20 млн видов животных (консументов и редуцентов). Следовательно, 1 вид сосудистых растений репрезентативно обуславливает пространственное распределение и жизнедеятельность 66 видов животных, грибов, бактерий и т.п.
Еще В.И. Вернадский отмечал «мозаичность» в строении биосферы, выделяя зоны «сгущения жизни». Глобально выделяется 3 географические области высокой биопродуктивности: широтные в пределах 35-40° северной и южной широт; прибрежно-морская зона мира протяженностью >700 тыс. км (содержащая 90% биопродуктивности Мирового океана); широтная зона так называемого Южного океана, окружающего Антарктиду, которая характеризуется сложныи взаимодействием глубинных и поверхностных вод – апвеллингами, даунвеллингами, гидрофронтами и т.п.
На рис. 7.9, отражающем общие закономерности глобального биологического разнообразия, четко проявлены его главнейшие особенности: широтная климатическая зональность и области (и участки) центров максимумов биоразнообразия, обусловленных их специфическими географическими условиями и историей их развития, включая рефугиумы (убежища видов).
Выделяется 6 центров максимального глобального разнообразия, каждый их которых содержит более 5000 видов растений на 10000 км2: Чоко (Коста-Рика), тропические Восточные Анды, приатлантическая Бразилия, Восточные Гималаи (провинция Китая Юннань), северный Борнео, Новая Гвинея.
Кроме того, выделяются 16 центров высокого биоразнообразия, каждый из которых содержит 3000 видов и более на 10000 км2, с пятнами наивысшего разнообразия.
Таково современное распределение этого важнейшего экологического фактора. Но в настоящее время, вледствии в основном антропогенного воздействия, под угрозой уничтожения находится более 30000 видов жи вотных и растений. Темпы исчезновения видов млекопитающих за последнее столетие возросли в 40 раз по сравнению с прошлыми периодами. За последние 400 лет исчезли 484 вида животных и 654 вида растений. А они имеют не только экономическую ценность, но являются необходимым условием существования жизни на Земле. Основными причинами их исчезновения, помимо использования и потребления, были: искусствен ная
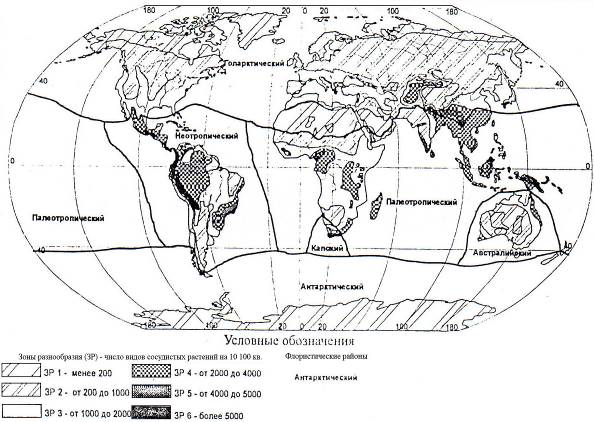
Рис.7.9. Схема биотического разнообразия суши мира
интродукция (39% от потерянных видов животных), разрушение условий существования (36%) и неконтролируемая охота (23%).
Основные факторы биологического разнообразия приведены на рис. 7.9.
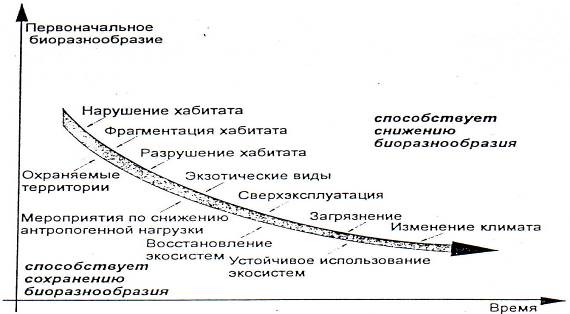
Рис. 7.10. Основные факторы изменения биологического разнообразия
Среди главных способов защиты биологического разнообразия два стратегических: in situ (в месте обитания) и ex situ (за их пределами). Главное направление стратегии должно быть на уровне экосистем, но не исключительно. Поэтому так необходимо особое внимание к организации и деятельности особо охраняемых природных территорий. Эрозия биологического разнообразия продолжается.
Гл. 8. ПАЛЕОЭКОЛОГИЯ
1. Предмет палеоэкологии
Палеоэкология (палеогеоэкология) как наука находится в стадии становления. Необходимость изучения древних экологических событий Земли для познания ее современных экологических особеностей несомненна. Возможности самоочищения природных сред относительно ограничены и в известной степени регламентируются интенсивностью тех природных загрязнений, которые зарегистрированы в летописях Земли, включая катастрофические. Именно ими определяются апробированные природой предельно допустимые нагрузки для соответствующих геосфер. Так как ряд антропогенных процессов загрязнения по своим масштабам стал сравним с естественными глобальными загрязнениями, - все, что превышает указанные допуски, переходит в разряд непредсказуемых по последствиям планетарных явлений, которые вероятностно ведут к крупным и глобальным катастрофическим явлениям.
Так как реагирование экосистем на внешние раздражители во многом определяется аппаратом "памяти", для познания ее природы крайне важна проблема истории данной экосистемы, прослеживание эволюции которой во времени является основной задачей рассматриваемой науки.
Истоки палеоэкологии в этом понимании преимущественно связаны с геологическими исследованиями (стратиграфией, палеонтологией, палеовулканологией, палеоокеанологией, палеоклиматологией и т.п.). Нередко к палеоэкологическим относят и некоторые аспекты результатов археологических изысканий, представляющих собой предсовременные этапы экологии Земли.
В более традиционном понятии палеоэкология - раздел палеонтологии, изучающий взаимосвязи между организмами и средой их обитания, а также условия отмирания организмов, их захоронения и последующую историю в геологическом прошлом на основе исследований окаменелых остатков фауны и флоры и их стратиграфического положения. Синонимом палеоэкологии является экологическая палеонтология - ее отрасль, изучающая условия существования и образ жизни организмов в прошлые геологические эпохи, а также соотношение и зависимость между организмами геологического прошлого и средой их обитания.
Таким образом, и здесь, как и в современной экологии, как в фокусе сошлись интересы наук о Земле и биологов.
На пути восстановления основных этапов развития жизни на Земле и самой Земли существуют кардинальные объективные трудности, во многом обусловленные современным и прогнозируемым уровнями познания.
Несмотря на попытки обобщения обильного нового материала, все еще крайне слабо распространены балансовые представления: чаще рассматриваются действия одного или очень немногих "ведущих" факторов. Эволюция косных систем определяется их начальным состоянием и воздействиями окружающей среды. В биологической эволюции жестко задано лишь общее направление - от простого к сложному, от меньшей устойчивости к большей, от менее замкнутых биогеохимических круговоротов к более замкнутым. Сама эволюция включает бесконечное число почти равновероятностных выборов и последствия их могут быть колоссальными. Восстановление систем всегда занимало значительно больше времени, чем их разрушение.
Древние экосистемы определенно существовали в двух различных состояниях: "холодном", как сейчас, и "теплом", на протяжении большей части геологической истории. Поэтому должны быть вычленены фундаментальные принципы организации и функционирования тех и других.
Характер биосферы в истории развития Земли оказывался существенно отличным из-за различной геологической основы соответствующего времени. Выделяются периоды относительно устойчивого состояния, когда материки не так расчленены и низки, а океаны мелководны - развиты эпиконтинентальные морские обстановки. Следовательно, и циркуляции воздушных и водных масс, а также особенности осадконакопления и другие должны быть существенно различны.
Древние экосистемы, как и современные, реагируют на неблагоприятные воздействия снижением многообразия. Поэтому изучение изменения разнообразия - один из важнейших инструментов в палеоэкологии. Лучше изучено, в силу большей палеонтологической информативности, биологическое разнообразие морских и континентальных водоемов; значительно менее - наземных экосистем.
В отличие от морских, в пресноводных и наземных экосистемах равновесные участки эволюции разнообразия - редкое исключение. Причина - в большем экосистемном единстве морских условий. В море экосистемы как бы нанизаны на единый поток вещества и энергии и не имеют возможности кондиционировать среду своего обитания. В результате все морские экосистемы в значительной степени реагируют как некое единое целое. Морские системы долго сопротивляются и сильно разрушаются, наземные и пресноводные изменяются по частям и не столь глубоко.
Географические параметры накладывают соответствующий отпечаток и на климатические особенности этих обстановок. Однако, бо-льшинство климатических изменений сами по себе приводят обычно не к принципиальным качественным переменам в биосфере, а к перераспределению площадей, занятых различными биомами.
Ритмика биосферы задается космическими и планетарными воздействиями, синхронизирующими темп эволюционных перестроек.
"Длительность периодов собственного времени биосферы затрудняет мониторинг экосистем, фиксируемые при нем изменения представляют собой сложную мозаику разнокачественных процессов, где на направление изменения наложен спектр колебаний" (Пономаренко, Розанов, с. 273).
Закон необходимого разнообразия У. Эшби гласит: разнообразию неблагоприятных внешних воздействий для сохранения устойчивости система может противопоставить только не меньшее разнообразие сохраняющих реакций.
Рамки данной работы не позволяют рассмотреть все разнообразие аспектов затронутой проблемы, и заставляют сосредоточиться лишь на некоторых наиболее важных особенностях развития биоты Земли.
Поскольку речь пойдет о важнейших событиях органической жизни геологического прошлого, давайте вспомним об основной канве эволюции биосферы.
2. Эволюция биосферы
Развитие органического мира Земли охватывает большую часть ее геологической истории. Предположительно автотрофные растения возникли 3-3,5 млрд. лет назад. О существовании организмов архейского времени имеются преимущественно косвенные данные. В протерозое найдены остатки водных синезеленых водорослей, в верхнем протерозое - многоклеточных (кишечнополостные и др.).
В кембрийский период существовали сотни видов трилобитов, брахиоподы, ракообразные, моллюски, медузы и пр. (рис. 8.1). Были распространены и разнообразные водоросли. В ордовике-силуре количество безпозвоночных возросло. Вместе с трилобитами выявлено множество членистоногих, в том числе раков и скорпионов. Именно членистоногии были первыми поселенцами суши. Появляются примитивные бесчелюстные рыбы - первые позвоночные. К этому же времени относится возникновение растительного покрова суши, включая псилофиты. Но можно высказать предположение об опережении наземной растительности, учитывая трофные цепи первопроходцев суши. Однако, пока это не доказано.
В девоне произошли значительные изменения в составе органического мира. Появились кистеперые рыбы и произошедшие от них амфибии. К концу периода относится появление первых насекомых. Растения псилофитового типа заменяются хвощами, папоротниками и голосеменными, достигнув расцвета в каменоугольный период.
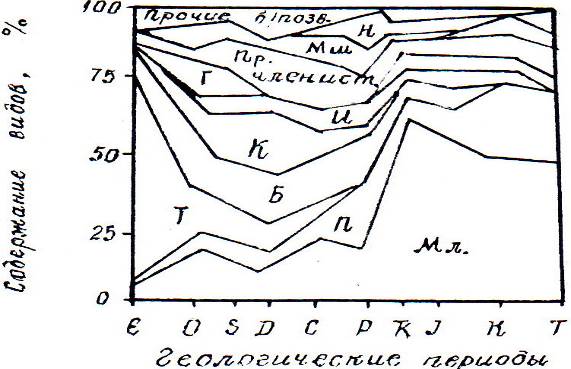
Рис. 8.1. Распространение животного мира в геологической летописи
Б – брахиоподы, Г – граптолиты, И – иглокожие, К – кишечнополостные, Мл – моллюски, Мш – мшанки, Н – насекомые, П – пресмыкающие, Пр – простейшие, Прочие б/позв. – беспозвоночные, Т – трилобиты
Наземные насекомые в карбоне достигают гигантских размеров.
К концу периода появляются рептилии.
В перми голосеменные (саговниковые и гингковые) приобретают доминирующее развитие. Процветали амфибии, прогрессировали ре-птилии.
Значительно обновился мир на рубеже мезозойской эры. Вымирают многие беспозвоночные, включая трилобитов, другие резко уменьшаются в численности.
С триаса открывается эпоха расцвета рептилий, продолжавшаяся весь мезозой. Возникли динозавры, черепахи, морские рептилии - ихтиозавры и плезиозавры. К концу триаса появляются первые примитивные млекопитающие.
В юрском периоде вымирают гигантские амфибии, но развиваются еще более огромные рептилии, включая разнообразных диназавров. Распространяются преродактили - летающие рептилии. Наряду с ними появляются первые птицы, во многом близкие рептилиям (археоптерикс - археорнис). В морях вместе с древними рыбами - акулами и скатами - существуют первые костистые рыбы. Млекопитающие все еще слабо распространены.
В меловом периоде появляются змеи, новые формы динозавров и крупные летающие ящеры - птеродонты.
Большую часть мезозоя на суше преобладают хвойные растения и папоротники. В середине мела появляются покрытосеменные, образовавшие широколиственные леса из дуба, тополя, бука, пальм и др.
Кайнозойская эра - новый важный рубеж в истории органического мира. К этому времени вымирает большинство рептилий, включая наземных динозавров, морских ящеров, летающих ящеров. Из разнообразных рептилий мезозоя сохраняются лишь крокодилы, черепахи, ящерицы и змеи.
В палеогене ускоряется эволюция плацентарных млекопитающих, появившихся в меловом периоде. Экологические ниши, освобожденные рептилиями, заняли млекопитающие, многие из которых обладали гигантскими размерами. Одни заселяли океан, другие обрели способность летать. Большое место в фауне кайнозоя заняли птицы.
Растительность в кайнозое мало изменилась, но лучше проявилась зональность при колебаниях климата, особенно в четвертичном периоде.
Конец кайнозоя завершается возникновением человека. До этого с палеоцена его эволюция была мало заметной. В нижнем олигоцене появляются первые человекообразные обезьяны, в плиоцене - австралопитеки, а затем и первые ископаемые люди. Современный человек появился в эпоху последнего оледенения, 35-40 тыс. лет назад.
Не следует полагать, что масса органического вещества постоянно росла. Если рассматривать ископаемые каменные угли, как отражение былой лесистости, при допущении примерно равной их сохранности в течении постдевонской истории Земли, то, с использованием шкалы абсолютного возраста, следует прийти к выводу о тенденции уменьшения объема растительности с позднего девона до наших дней; при отдельных вспышках и угасаниях. Вероятно, что от этого зависела и масса животных.
В целом "древо" биологической эволюции может быть обобщено в виде следующей схемы (рис. 8.2).
3. Важнейшие биотические кризисы
Следует иметь в виду и то, что время в геологии обладает характеристикой "спрессованности": чем древнее "каменные летописи", тем больше в них заключено временного диапазона при равных мощностях вмещающих фауну и флору отложений.
Рамки "оживающего" геологического времени все время раздвигаются, а появление первого живого существа все более отодвигается в глубь времен. Многие разночтения в этом отношении определяются степенью сохранности ископаемых органических остатков. В общем случае лучше сохраняются скелетные образования, хуже (или не сохраняются) ткани. Да и сами находки достаточно разнолики, не всегда даже ясен вопрос об их органичности (так называемая палеонтологическая проблематика). Зависит степень изученности и от технически-приборного уровня исследований.
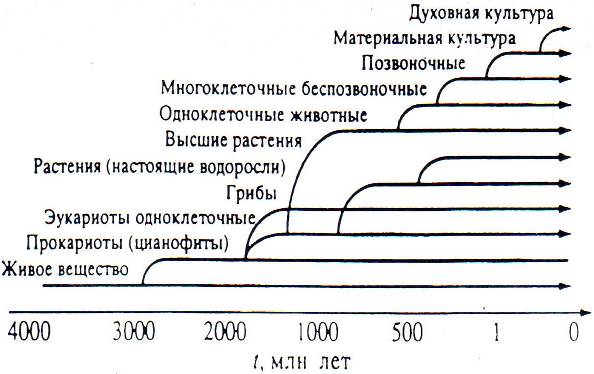
Рис. 8.2. "Древо" биологической эволюции
"Совсем недавно, всего два-три десятка лет тому назад, мысль о том, что микробы могут сохраняться в ископаемом состоянии, особенно в массовых количествах, могла бы показаться абсурдной" (Розанов, 1999, с.63). А уже считается, что карбонатные микриты (очень мелкозернистые породы) в своей массе это результат деятельности бактерий. В основе лежат уникальные исследования по искусственной фоссиликации (литофикации) ныне живущих цианобактерий. Чехлы нитей их окаменевают за несколько часов. Так зародилась "бактериальная палеонтология", утверждающая ныне, что, за редким исключением, все осадочные породы в той или иной степени образовались с участием микробов.
Еще несколько лет назад считалось, что первые многоклеточные появились в венде (ок. 650-550 млн лет назад), а одноклеточные эукариоты около 1,2-1,3 млрд. лет тому назад. Теперь мы знаем о находках низших грибов в породах, возраст которых примерно 2,2 млрд., а первых многоклеточных - около 1,4 млрд. лет тому назад.
В 1969 г. появилась молекулярная палеонтология (Кельвин). В последние 10 лет вышли работы по изучению ДНК у вымерших форм. Получены копии участков генов из головного мозга первобытного человека. ДНК выделена из остатков мамонтов (10-50 тыс. лет назад).
Удивительный результат получен И.С. Барсковым, обнаружившим фосфатизированные кровеносные сосуды у позднеюрского аммонита из района Воробьевых гор в Москве. Проведено томографическое исследование двух черепов енкилозавров (панцерных динозавров), сканирование которых позволило воспроизвести внутренние структуры головы: носовые проходы, синусы, мозговую полость (Университет Пенсильвании). Обнаружена даже остеома.
Вместе с тем не надо обольщаться: вряд ли мы способны изучить и 1% бывшего богатства органического мира.
Поэтому наши современные взгляды не могут не быть глубоко ошибочными. Но такова судьба всякого научного поиска.
Например, сейчас многое базируется на известных нам к настоящему времени данных о скоростях эволюционных преобразований, о темпах возникновения таксонов, начиная с уровня классов (рис. 8.3).
Постоянное обновление органического мира в результате общей эволюции жизни на Земле и общий прогресс науки в этом направлении достаточно полно отражают две сравнительные диаграммы динамики вымираний семейств морских организмов в фанерозое (А - Дж. Филлипс, середина XIX в., и Б - Алексеев, 1989 г.) - рис. 8.4.
Из самого беглого сравнения очевидно, что общий характер современных научных достижений свидетельствует о том, что вымирания между этапами не были столь резки, обновление органического мира не было столь кардинальным. Но общее число вымираний известных нам достаточно велико, а более мелких флуктуаций очевидно бесконечное множество.
Пока анализу подаются лишь наиболее массовые вымирания, причины которых являются предметом многих научных дискуссий.
Кембро-ордовикский кризис отвечает временному интервалу, в течении которого мог осуществиться переход от ледникового состояния к среднепалеозойскому парниковому состоянию.
Ордовикско-силурийский кризис совпал с оледенением и соответствует резкому и кратковременному переходу к другим климатическим ус-
ловиям. Или же это действие неизвестного внешнего фактора, вызвавшего как оледенение, так и биотический кризис.
Позднедевонский кризис возможно был обязан переходу от среднепалеозойского парникового состояния к позднепалеозойскому ледниковому.
Кризис на границе перми и триаса и последней части позднепермско-триасового холодного интервала наступил после оледенений, но задолго до раскола Пангеи. Возможные причины его - резкое понижение уровня моря и возникновение солоноватых приповерхностных вод за счет различных факторов.
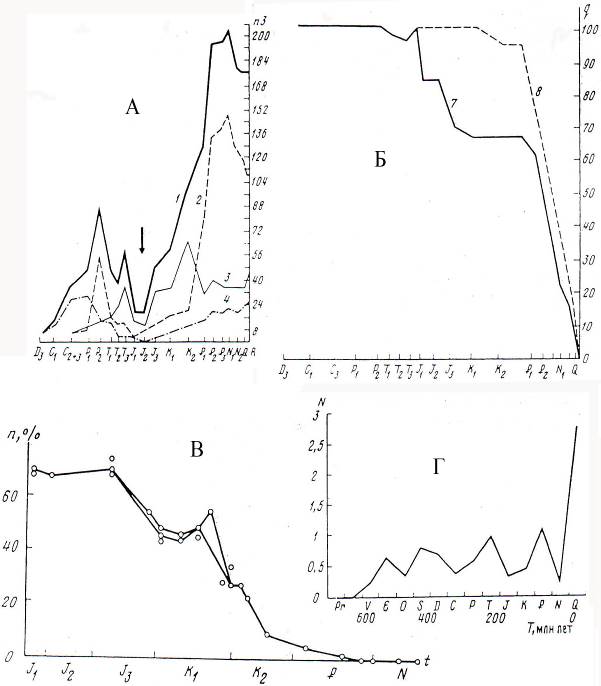
Рис. 8.3. А. Динамика таксонометрического разнообразия наземных
тетрапод (на уровне семейств)
Б. Доля вымерших семейств тетрапод
В. Изменение доли вымерших семейств насекомых
Г. Изменение скорости (частоты) N (10 5-6 0 такс/лет)
возникновения таксонов (от класса и выше)
Триасово-юрский кризис фиксирует переходное климатическое состояние, возникшее вскоре после начала рифтообразования, вслед за которым наступили парниковые условия.
Кризис на границе мела-перми отличается от всех других своей внезапностью. Даже в областях непрерывного морского осадконакопления он может быть привязан к определенной поверхности напластования или к тонкому слою "пограничной глины". В ней наблюдаются высокие концентрации иридия. Относится ли к этому времени вымирание динозавров - не определено. Несомненно, что он не был связан с длительной парниковой инверсией, так как мезозойские парниковые условия продолжались до эоцена.
Кризис на границе верхнего эоцена-олигоцена ярко выражен в пелагической области: переход от мезозойско-эоценового парникового эффекта к последующему состоянию оледенения.
Таким образом, основные переходные моменты между главными климатическими эпизодами в истории Земли отвечают следующим кризисам: постпозднекембрийскому (ок. 500 млн лет назад), позднедевонскому (ок. 355 млн лет), концу триаса (192 млн лет), позднеэоценовому (ок. 40 млн лет назад). Интервалы между ними составляют 145, 163 и 152 млн лет.
Из всех перечисленных кризисов - явлений, когда стресс вызывает замену главных структур системы, а последняя сохраняется в связи с поглощением этого стресса подсистемами и происходит замена старой системы новой - два претендуют на оценку близкую к катаклизмам - полному разрушению системы и подсистем. Это глобальный "фанерозойский кризис" Мак-Алистера (конец перми) и географически более ограниченный региональный - "мессинский" (позднемиоценовый).
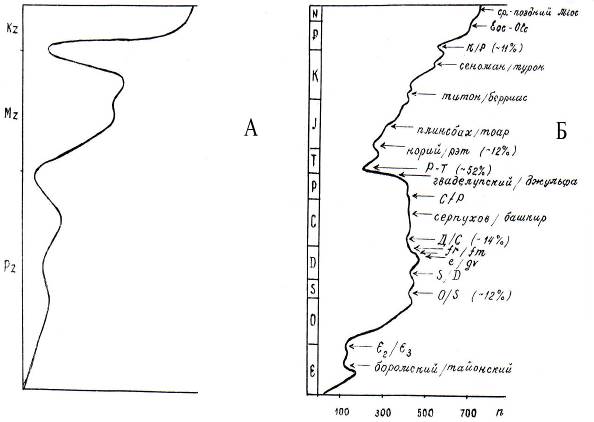
Рис. 8.4. Динамика вымираний семейств морских организмов в фанерозое
За время "фанерозойского кризиса" разнообразие животных семейств и в море и на суше резко упало с 228 до 161. Этот мор пытаются объяснить отравлением организмов рассеянными элементами, сокращением площадей обитания, радиацией, изменением пищевых ресурсов, высокими температурами и колебаниями солености вод. Чаще всего его связывают с увеличением солености морских вод.
Предполагают отток рассолов и формирование глубоководного слоя высокой солености (подобно современному Средиземному морю). В верхних слоях моря соленость снизилась с 35%о до 30%о в результате удаления (осаждения) около 3 млн км3 растворенных солей (в пермских отложениях их находится ок. 1,59 млн км3). Этот период соответствует начальному этапу образования океана Тетис, который закрылся в миоцене (5-6 млн лет назад).
Может быть предпочтительней другое объяснение: возможное сокращение областей обитания на шельфах.
Наоборот, в "мессинский кризис солености" происходит финальное событие в развитии океана Тетис в позднем миоцене, когда прервалась его последняя связь с Мировым океаном (орогенные комплексы Бетико и Рифа закрыли Иберийский проход). Океан превратился в глубокое теплое море, уровень воды в котором понизился на 70 м, а эвапориты в цепи озер сконцентрировались в объеме 1,2 млн км3, что повсюду вызвало вымирание стеногалинной морской фауны. При переходе от миоцена к плиоцену сменилось более 40% видов морских организмов.
Кризис закончился внезапно разрушением преграды, отделявшей озера-моря от Атлантического океана. Вновь возникло Средиземное море.
Одна из моделей этого события обобщает следующую главную цепь причин.
Изменчивые параметры земной орбиты
влияют как на инсоляцию, так и на
Регрессия область перехода ядра и мантии. Из-
меняются геомагнитное и гравитаци-
онное поля
Понижение геоида под континентами
приводит к соответствующему пониже-
нию уровня грунтовых вод
Засуха
Смена растительности
Фаунистические изменения
Вымирание
Выделяются два главных суперцикла фанерозойской истории, отраженные в соответствующих изменениях уровня Мирового океана (рис. 8.5).
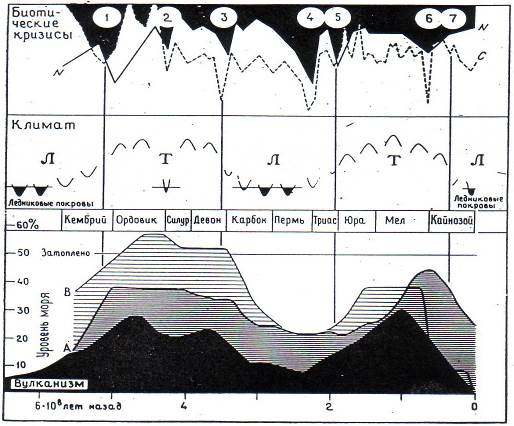
Рис. 8.5. Два суперцикла фанерозойской истории
Уровни моря: А - по Вейлу и др. Б - по Хэллему
4. Модели вымирания
Из всех вымираний наиболее полно изучено массовое вымирание в конце мелового периода. Оно коснулось и растительности и животного мира. Однако, все имеющиеся данные говорят не о всеобщей катастрофе, а о сложном характере вымирания, которое продолжалось миллионы лет.
"Отвратительная тайна" Ч. Дарвина - покрытосеменные - находились уже в середине мощной эволюционной экспансии и стали доминировать в наземной флоре только четверть периода назад (рис. 8.6).
Главными тенденциями в развитии покрытосеменных были усиление провинциализма и постепенное возникновение новых форм, принадлежащих к современным семействам и даже родам.
За последние годы был предложен ряд впечатляющих "сценариев" биотического вымирания в конце мелового периода.
Альварес предположил столкновение с астероидом ориентировочно размером 6-14 км в диаметре, которое привело к огромному об лаку пыли и остановке на ряд лет фотосинтеза. Он же допускает фрикционный нагрев атмосферы.
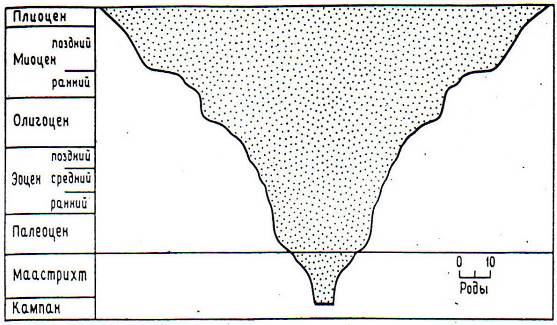
Рис. 8.6. Рост числа современных родов покрытосеменных
Ксю обратился к столкновению Земли и кометы с возможным отравлением цианидом из головной части последней.
Выдвигались гипотезы вспышки сверхновых звезд или Солнца, высоких радиаций.
Но явно, что происходило и временное похолодание климата (рис. 8.7).
Палинологические данные (пыльца и споры) - самые надежные свидетельства смены наземной растительной биоты на границе мелового и палеогенового периодов. Однако, обычно данные эти ограничены (средняя коллекция пыльцы может содержать 800-1000 видов, а для стратиграфических целей используется только 200). Но разового исчезновения (элиминации) также не отмечается. Поэтому нет убедительных палинологических данных, подтверждающих идею всемирной катастрофы. В самом конце мелового периода и в палеогене наземная флора менялась главным образом за счет взрывной эволюции и диверсикации различных видов и соответствующего уменьшения относительного содержания голосеменных, папортников и близких к ним групп. Наблюдается постепенный флористический переход. Изменения эти не более сильные, чем те, которые могли наблюдать на границах эпох или даже внутри подотделов эпох. Причины, вероятно, следует искать в климатических колебаниях, возникших в результате перемещений континентальных плит или спрединга океанического дна.
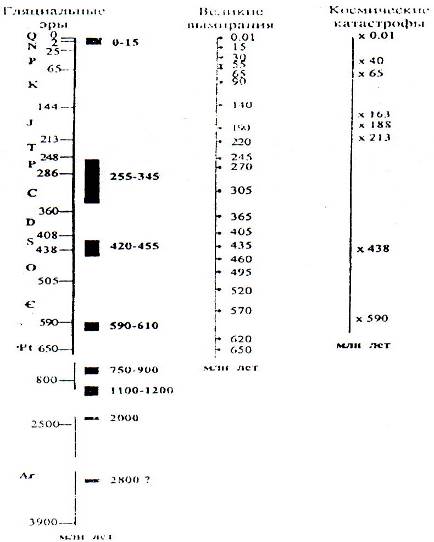
Рис. 8.7. Крупнейшие стрессы в истории биосферы
Нельзя прийти к выводам о катастрофе и по анализу млекопитающих.
Вымирание же крупных рептилий действительно «не предвещало ничто», но оно фактически произошло. Как и многих других групп морских организмов. Мы лишь знаем, что терминальное (конечное) меловое вымирание в морских условиях продолжалось от 1 до 5 млн лет. Несомненно, что первоначально оно явилось следствием общего ухудшения экологической обстановки: крупномасштабных изменений уровня моря, химического состава воды (особенно соединений кислорода), температуры и циркуляции воды в океане, климата, общим разрушением экологических комплексов. На последней фазе упадка биоты ее вымирание было усилено каким-то событием внеземного происхождения.
Более ясна модель меловых вымираний. Она выглядит примерно следующим образом:
1. Крупный эвстатический подъем уровня моря (в связи с поднятием хребтов на дне из-за активного движения плит) сопровождается глобальной трансгрессией, улучшением и потеплением климата и увеличением разнообразия экологических условий.
2. Увеличивается разнообразие морских фаун, темп эволюции.
Смешиваются умеренно теплолюбивые, тетические и космополитные биоты. Повсеместная конкуренция за пищу и пространство в зоне смешения нарушает первичную структуру экосистем, улучшение климата ведет к снижению температурных градиентов в Мировом океане. Это благоприятствует увеличению разнообразия стенотермных организмов. Фактически большие части морской экосистемы становятся "подготовленными к гибели".
3. Быстрое изменение одного или нескольких экологических факторов наносит сильный удар по экосистеме в тот период, когда разнообразие тепловодных групп очень высоко и идет перестройка крупных экологических комплексов. Тепловодные таксоны вымирают. Морская экосистема разрушается. Причиной таких событий видимо было глобальное понижение содержания кислорода в Мировом океане. Само это бескислородное событие явилось результатом понижения термального градиента на поверхности океана из-за поднятия уровня моря, повышения температуры и вероятно солености; в конечном счете, нарушении водной циркуляции. Результатом этого "удара" были волны вымирания: сначала бентосной микрофауны и стенобионтной эндобентостной макрофауны, а затем эпибентосной макрофауны и аммонитов. Такой удар по среде обитания происходит за 0,5-2,0 млн лет до окончательного вымирания данной биоты.
4. Одновременно с первоначальным стрессом (или несколько опаздывая), накладывается несколько экологических факторов (быстрое изменение температуры, эвстатических понижений, потеря мелководных биотопов). Возникают условия, при которых возможна полная гибель крупных частей экосистемы. Именно величина биологического стресса, возникшего в результате многофакторного ухудшения среды, является самой главной причиной крупных вымираний.
В этой фазе происходит окончательное вымирание уже угнетенных тепловодных макрофаун, включая биоту мелководий. Наступает спад у бентосной макрофауны шельфов более умеренной зоны, некоторых групп пелагических микробиот и роющей ихтиофауны.
Стресс испытывает вся морская система, и впервые он затрагивает самые мелководные и верхние пелагические сообщества. Оказывается в тисках стресса и стенотермная биота. Понижение содержания кислорода вызывает миграцию в верх зоны кислородного минимума, которая достигает шельфов и т.п.
Следовательно, драматическое маахстрихское вымирание экологически различных планктонных организмов и тропического макробентоса может быть объяснено стрессом их общего биотопа - верхних слоев воды тропических-субтропических океанов.
Главный этап вымирания тетической пелагической микробиоты наступает после вымирания мелководной тетической макрофауны.
5. Окончательный кризис биоты совпадает с пиком ухудшения среды, обусловленной многими факторами. В последнюю очередь гибель затрагивает пелагический микропланктон. Гибель известкового планктона была "катастрофической" - за несколько десятков тысяч лет. Сократилось первое звено пищевой цепи.
Последнее крупное бескислородное событие выразилось в широком распространении зоны кислородного минимума на верхние уровни водной толщи. Ему соответствуют богатые органическим веществом "пограничные глины", "рыбные слои" (массовая гибель ?), ограничение биотурбации и сокращение бентосной биоты.
Близкие по своей сути к описанным этапы кризисов выделяются В.В. Жерихиным и А.С. Раутианом под названиями предкризиса (подготовительные фазы) и собственно кризиса, состоящего из параксимальной и драматической фаз. Кроме этого выделяется посткризис (фаза успокоения).
Однако, существуют и другие механизмы быстрого фаунистического переворота, отраженные в гипотезе равновесия Мак-Артура-Уилсона (рис. 8.8).
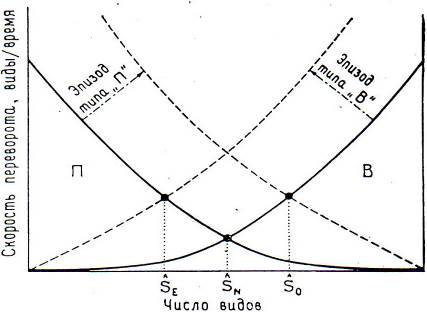
Рис. 8.8. Скорость фаунистических переворотов
В - вымирание таксонов, П - появление таксонов
Для вымирания таксонов (В) толчком служат резкие изменения обстановки, в результате чего число таксонов уменьшается; для появления таксонов (эпизоды типа П) характерен большой приток иммигрантов, приводящий к увеличению числа таксонов.
Пример эпизода В: В Северной Америке коарендонтская хронофауна (разнообразная, преимущественно копытная фауна саванн), которая достигла расцвета между 15 и 18 млн лет назад, была уничтожена в течении нескольких следующих млн лет в результате ухудшения климата и перерождения в скудную степную фауну. Возможные причины: региональные поднятия и мессинское и климатические события, произошедшие около 5 млн лет назад. Восстановление фауны заняло 2 млн лет.
Пример эпизода П: Южно-Американская фауна стала заметно богаче за счет пришедших с севера во время Великого Американского переселения иммигрантов (3 млн лет назад). Число родов увеличилось на 30-50%. И не падало до прежнего уровня.
Но эти эпизоды могли и взаимодействовать.
Если за эпизодом В последует П (В-П) - вначале появляются крупные экологические ниши, которые интенсивно заполняются. Вероятно, великие революции в истории развития жизни представляют собой такие сложные эпизоды (в отличие от великих вымираний).
Развитие работ по изучению исторических закономерностей эволюции биосферы (а это возможно только на палеонтологическом материале) ясно показало, что биосфера является хорошо сбалансированной, циклично развивающейся авторегулируемой системой, которая пережила все внешние возмущения за 4,5 млрд. лет существования Земли. Стало очевидно, что только изучение естественных трендов эволюции компонентов биосферы и симптомов кризисов прошлого может дать реальное представление об экологической стратегии поведения человека в условиях антропогенного кризиса.
Дата добавления: 2016-01-07; просмотров: 1359;